
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Дипломная работа: Изучение влияния селенсодержащих соединений на системную гемодинамику и мозговой кровоток
Дипломная работа: Изучение влияния селенсодержащих соединений на системную гемодинамику и мозговой кровоток
СОДЕРЖАНИЕ
ВВЕДЕНИЕ
ГЛАВА 1 ОБЗОР ЛИТЕРАТУРЫ
1.1 Современные представления о механизмах регуляции мозгового кровотока
1.2 Роль селена в организме
1.3 Фармакологическая характеристика селенсодержащих соединений
ГЛАВА 2 МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
2.1 Изучение острой токсичности
2.2 Регистрация мозгового кровотока методом водородного клиренса
2.3 Регистрации параметров кардиогемодинамики с помощью компью-терной программы «Bioshell» на бодрствующих животных
2.4 Методика оценки поведения животного в тесте «открытое поле»
2.5 Оценка координации движений
2.6 Методика условного рефлекса пассивного избегания (УРПИ)
2.7 Методы изучения противогипоксической активности
2.8 Методика воспроизведения постишемических цереброваскулярных феноменов
2.9 Условия экспериментальных исследований
2.10 Статистическая обработка данных
ГЛАВА 3 ВЛИЯНИЕ СЕЛЕНИТА НАТРИЯ И СЕЛЕНИТА ЦИНКА НА ЦЕРЕБРАЛЬНУЮ ГЕМОДИНАМИКУ, СИСТЕМНОЕ АРТЕРИАЛЬНОЕ ДАВЛЕНИЕ И ЧАСТОТУ СЕРДЕЧНЫХ СОКРАЩЕНИЙ УСЛОВИЯХ ЭКСПЕРИМЕНТАЛЬНОЙ НОРМЫ
3.1 Изучение острой токсичности исследуемых соединений
3.2 Влияние селенита натрия и селенита цинка на мозговой кровоток
3.3 Влияние селенита натрия и селенита цинка на артериальное давление и частоту сердечных сокращений у бодрствующих крыс
ГЛАВА 4 ВЛИЯНИЕ СЕЛЕНИТА НАТРИЯ И СЕЛЕНИТА ЦИНКА НА ПСИХОНЕВРОЛОГИЧЕСКИЙ СТАТУС ЖИВОТНЫХ В УСЛОВИЯХ ЭКСПЕРИМЕНТАЛЬНОЙ НОРМЫ
4.1 Изучение влияния селенита натрия и селенита цинка на познавательную, двигательную и эмоциональную активность
4.2 Изучение влияния исследуемых соединений на процессы памяти и обучения
4.3 Влияние селенита натрия и селенита цинка на координацию движений
ГЛАВА 5 ИЗУЧЕНИЕ ПРОТИВОГИПОКСИЧЕСКОЙ АКТИВНОСТИ ИССЛЕДУЕМЫХ СОЕДИНЕНИЙ И ИХ ЭФФЕКТИВНОСТИ В УСЛОВИЯХ ЭКСПЕРИМЕНТАЛЬНЫХ ЦЕРЕБРОВАСКУЛЯРНЫХ РАССТРОЙСТВ
5.1 Влияние селенита натрия и селенита цинка на устойчивость животных к гипоксической гипоксии
5.2 Влияние селенита натрия и селенита цинка на устойчивость животных к циркуляторной гипоксии мозга, вызванной билатеральной окклюзией общих сонных артерий
5.3 Влияние селенита натрия и селенита цинка на устойчивость животных к тотальной ишемии мозга, вызванной критическими гравитационными перегрузками
5.4 Влияние исследуемых соединений на мозговой кровоток у наркотизированных белых крыс в условиях глобальной ишемии мозга
5.5 Влияние селенита натрия и селенита цинка на артериальное давление и сопротивление мозговых сосудов в условиях экспериментальной патологии
ГЛАВА 6 ИЗУЧЕНИЕ ПСИХОТРОПНОЙ АКТИВНОСТИ СЕЛЕНИТА НАТРИЯ И СЕЛЕНИТА ЦИНКА В УСЛОВИЯХ ТОТАЛЬНОЙ ИШЕМИИ МОЗГА
6.1 Изучение влияния селенита натрия и селенита цинка на психоневрологический статус белых крыс, подвергшихся воздействию гравитационных перегрузок (профилактическое введение)
6.2 Изучение терапевтического действия исследуемых соединений на психоневрологический статус животных, подвергшихся воздействию гравитационных перегрузок
ЗАКЛЮЧЕНИЕ
ОБЩИЕ ВЫВОДЫ
СПИСОК ЛИТЕРАТУРЫ
СПИСОК ИСПОЛЬЗУЕМЫХ СОКРАЩЕНИЙ
GPX (ГлП) - глутатионпероксидаза
АД – артериальное давление
АХ - ацетилхолин
ГАМК - гамма-аминомаслянная кислота
МК – мозговой кровоток
НМК – нарушения мозгового кровообращения
НТ – нейротрансмиттеры
САД – системное артериальное давление
СБ - селенсодержащие белки
СТС - сигнал-трансдукторные системы
ЦНС – центральная нервная система
ЧСС – частота сердечных сокращений
КА - катехоламины
Несмотря на достигнутые успехи практической кардиологии в борьбе с возникновением и последствиями сердечно-сосудистой патологии проблема фармакологической коррекции кардиологических заболеваний остаётся одной из актуальных проблем медицины. В структуре сердечно-сосудистых заболеваний одно из ведущих мест занимают сосудистые поражения мозга [1].
В последнее время в практике лечения больных с нарушениями мозгового кровообращения получили широкое распространение антиоксиданты. Применение антиоксидантов используется в терапии ишемического инсульта с целью оптимизации окислительно-востановительных процессов. В проведённых экспериментальных и клинических исследованиях высокую антиоксидантную активность показал селенит натрия [2].
По данным С.М. Николаева селенит натрия предохраняет миокардиальные клетки от разрушения, ограничивает периинфарктную зону, уменьшает размеры рубцов [3]. М.Д. Савиной и А.Н. Кудриным выявлен антиаритмический эффект селенита натрия на экспериментальных моделях аритмий [4]. Накоплен большой опыт применения селенсодержащих соединений в качестве пищевых добавок [2].
Высокая эффективность селенита натрия связана с его способностью повышать активность GPX. Этот фермент способствует накоплению глутатиона, прекращающего или ограничивающего процессы ПОЛ и его распространение [5, 6, 7].
Однако действие селенсодержащих соединений на системное артериальное давление, мозговой кровоток до сих пор не изучено. Учитывая тот факт, что селениты являются выраженными антиоксидантами, представляло интерес изучить их влияние на показатели системной гемодинамики и оценить целесообразность применения в качестве нейропротекторов.
Цель работы: изучение влияния селенита натрия и селенита цинка на параметры системной и церебральной гемодинамики и психоневрологический статус животных в условиях нормы и экспериментальной патологии.
Задачи исследований:
1. Изучить влияние селенита натрия и селенита цинка на мозговой кровоток, системное артериальное давление и частоту сердечных сокращений.
2. Изучить влияние селенита натрия и селенита цинка на психоневрологический статус животных в условиях экспериментальной нормы:
- двигательную активность и эмоциональные реакции крыс в тесте «открытое поле»;
- память и обучаемость животных в методике пассивного избегания;
- координацию движений.
3. Изучить противогипоксическую активность исследуемых соединений.
4. Изучить влияние исследуемых соединений на показатели системной и церебральной гемодинамики в условиях глобальной ишемии мозга.
5. Изучить психотропную активность селенита натрия и селенита цинка в условиях тотальной ишемии мозга, вызванной критическими гравитационными перегрузками.
Научная новизна исследований
В результате проведённого исследования впервые получены данные, согласно которым селенит натрия и селенит цинка вызывают снижение мозгового кровотока у наркотизированных животных. Длительное профилактическое введение селенита натрия и селенита цинка вызывает снижение уровня эмоциональной тревожности. Селенит натрия и селенит цинка обладают выраженным антигипоксическим эффектом: повышают устойчивость животных к гипоксической и циркуляторной гипоксиям. В условиях острой сосудистой патологии мозга селенит натрия и селенит цинка снижают смертность животных, повышают двигательную и исследовательскую активность, уменьшают тревожность, улучшают процессы обучения и памяти, координацию движений. Предварительное введение селенита натрия и селенита цинка до ишемии мозга уменьшает степень проявления фазы гиперперфузии и тормозит развитие фазы невосстановления мозгового кровотока
Практическая значимость работы
Полученные в экспериментальных исследованиях данные представляют теоретический и практический интерес, поскольку раскрывают неизученные эффекты селенита натрия и селенита цинка на мозговое кровообращение и психоневрологический статус в условиях нормы и при патологических состояниях. В клиническом плане особое значение имеет выявленная нами способность селенита натрия и селенита цинка уменьшать фазу гиперперфузии и тормозить развитие постишемического цереброваскулярного феномена невосстановления мозгового кровотока. Оба соединения представляют несомненный интерес в качестве средств для коррекции церебральных ишемий – самого распространённого вида сосудистой патологии мозга. Полученные результаты об анксиолитическом и ноотропном эффектах исследуемых соединений дают обоснование для рекомендаций по их рациональному использованию в комплексном лечении нарушений мозгового кровообращения.
Внедрение результатов исследования в практику
Результаты исследования нашли отражение в информационном письме «Обоснование применения селенита натрия и селенита цинка в комплексном лечении нарушений мозгового кровообращения», и направлены для информирования врачей МУЗ ЦГБ г. Пятигорска, МУЗ ЦГБ «Городская больница №2» г. Пятигорска, поликлиники 1 г. Пятигорска, МУЗ КЦГБ г. Кисловодска.
4. В условиях острой сосудистой патологии селенит натрия и селенит цинка снижают смертность животных, повышают двигательную и исследовательскую активность, уменьшают тревожность, улучшают процессы обучения и памяти, координацию движений.
ГЛАВА 1 ОБЗОР ЛИТЕРАТУРЫ
1.1 Современные представления о механизмах регуляции мозгового кровотока
Регуляция мозгового кровообращения давно является актуальной проблемой. Несмотря на большой фактический материал, накопленный по данной проблеме, вопрос о физиологических механизмах лежащих в основе регулирования мозгового кровообращения остаётся открытым. Мозг человека, составляя лишь 2% от общей массы тела, утилизирует 20-25% потребляемого организмом кислорода. Кровообращение в головном мозге нельзя рассматривать изолированно от кровообращения всего организма в целом. В неразрывной связи с МК находятся такие параметры, как системное артериальное давление (САД), общее периферическое сопротивление сосудов, центральное венозное давление, объем циркулирующей крови и т.д. Кровоснабжение мозга характеризуется наличием оптимального режима, обеспечивающего в процессе жизнедеятельности непрерывное и своевременное пополнение его энергетических и иных затрат. Это достигается последовательным включением ряда факторов, приводящих в действие механизмы саморегуляции мозгового кровообращения [8, 9, 10, 11, 12].
Регуляция мозгового кровотока крайне сложна. Из литературы можно заключить, что в регуляции принимает участие несколько взаимосвязанных регуляторных контуров, каждый из которых может функционировать самостоятельно. Действие их направлено на поддержание в определенных пределах химического состава ткани мозга (а соответственно и оптимальных режимов питания) и регуляции физического статуса мозговой ткани (ее объема, количества жидкости и т.д.). Выделяют 4 регуляторных контура: нейрогенный, гуморальный, метаболический и миогенный [13, 14, 15].
Роль нервной системы
В настоящее время считается общепризнанным активное участие нервной системы в регуляции мозгового кровообращения. Результаты морфологических исследований показали наличие в стенках мозговых сосудов адренергической и холинэргической иннервации.
Симпатическую иннервацию сосуды головного мозга получают от узлов шейного отдела симпатического пограничного ствола, включая звездчатый ганглий [15, 16, 17, 18]. На различных животных было установлено, что раздражение шейных и позвоночных нервов вызывает сужение сосудов мозга и уменьшение мозгового кровотока. В ряде исследований показано важное значение клеток синего пятна, которое представляет собой основное норадренергическое образование мозга. В них обнаружено высокое содержание катехоламинов [19, 20].
Важное значение в регуляции МК имеет холинергическая система. Парасимпатические волокна отходят от лицевого нерва и коленчатого ганглия, присоединяясь к синокаротидному сплетению, проникают в полость черепа [9]. Холинергические сплетения обнаружены в адвентиции артерий и пиальных сосудов. Гистохимические исследования показали, что волокна симпатических и холинергических нервов следуют рядом, образуя единое терминальное сплетение [21, 22, 23, 24].
Роль парасимпатической системы в регуляции мозгового кровообращения неоднозначна. Исследования показали противоречивые результаты. При стимуляции парасимпатических нервов и введении ацетилхолина обнаружили расширение сосудов мозга, которое снималось атропином. Активное расширение пиальных артерий в ответ на снижение системного артериального давления устраняется внутривенным введением атропина. Тогда как стимуляция холинергических ветвей большого каменистого нерва не приводила к изменению тотального мозгового кровотока [25].
Особый интерес представляет изучение роли аортальных и каротидных рефлексогенных зон, содержащих баро- и хеморецепторы в регуляции мозгового кровообращения. В ряде исследований [26, 27, 28, 29] активация каротидных барорецепторов повышением давления в каротидном синусе или электрическая стимуляция синокаротидных нервов вызывали расширение сосудов мозга параллельно снижению артериального давления. В ответ на включение каротидных барорецепторов пережатием общей сонной артерии или денервации каротидных зон – сосуды суживались, и артериальное давление повышалось. Другие исследователи [30] наблюдали противоположные реакции сосудов. Наряду с холинергической иннервацией мозговых сосудов современные исследователи обнаружили наличие пуринергических и пептидергических нервных волокон в стенках мозговых сосудов [31]. В артериях головного мозга найдены моноаминоациты (тучные, хромафинные клетки, меланофоры), которые функционально связаны в единую систему биологических регуляторов [32].
Исследования последних лет [33] подтверждают важную роль ствола мозга в обеспечении адекватного церебрального кровотока. Показаны нарушения регуляции кровотока в эксперименте при разрушении стволовых и спинальных путей [34]. Роль нервного компонента в местных сосудистых реакциях головного мозга была показана Ю.А. Москаленко и его сотрудниками [13, 26, 35, 36, 37].
В настоящее время была обобщена большая группа химических передатчиков сигналов между нейронами и от нейронов на эффекторные клетки получившая название нейротрансмиттеры (НТ). Именно НТ создают возможность объединения отдельных нейронов в целостный головной мозг и позволяет ему успешно выполнять все его многообразные и жизненонеобходимые функции.
Нейротрансмиттеры делят на нейромедиаторы – прямые передатчики нервного импульса, дающие пусковые эффекты (изменение активности нейрона, сокращение мышцы, секрецию железы), и нейромодуляторы – вещества модифицирующие эффект нейромедиаторов. Соотношение концентрации и активности нейромедиаторов определяет функциональное состояние большинства постсинаптических клеток. Нейромодуляторы обычно действуют более локально в определённых зонах мозга [38, 39].
Большинство НТ синтезируется в нейронах. Затем они транспортируются в везикулы в обмен на накопленные там Н+ (аккумуляция протонов в везикулах осуществляется особой Н+-АТФазой за счёт энергии АТФ). Эти везикулы расположены в нервном окончании, НТ хранятся в них в высоких концентрациях (до 100-500 мМ). Когда распространяющийся по нерву потенциал действия приходит в зону везикул, он открывает потенциалзависимые Са2+-каналы, ионы Са2+ входят в нервные клетки, что приводит к выбросу их из НТ в синапс.
Различают возбуждающие и ингибирующие, или тормозящие НТ. Эффекты первых преобладают в состоянии бодрствования и высокой функциональной активности, вторых - в покое и особенно во время спокойного сна без сновидений. По химической структуре НТ можно разделить на 5 классов: аминокислоты, амины и их производные, нейропептиды, нуклеозиды и нуклеотиды, стероиды. Последние 2 класса пока представлены едиными веществами [40, 41].
Все НТ диффундируют через синапс и на наружной стороне плазматической мембраны постсинаптической клетки связываются со специфическими рецепторами. НТ не требуется проникать через мембрану клетки, внутрь клетки поступает не сам НТ, а сигнал, возникающий при связывании НТ с рецептором. Восприятие, преобразование, усиление и передачу сигнала внутрь клетки и затем внутрь её органелл осуществляют сигнал-трансдукторные системы (СТС). Рецепторами нейромедиаторов являются регуляторные субединицы быстрых ионных каналов (Na+ – или Сl-) это ионотропные рецепторы. Эффекты нейромодуляторов реализуются более сложными СТС, включающими рецепторы, ГТФ-зависимые G-белки, мембранные ферменты, Ca2+ - или К+ - каналы, вторые посредники и их белковые рецепторы (чаще всего протеинкиназы) это метаботропные рецепторы. Разные механизмы реализации сигналов определяют временные различия: нейромедиаторы действуют за время нервного импульса миллисекунды (быстрые ответы клеток), модуляторы – за секунды или минуты, такие эффекты называются медленными.
Главные медиаторы головного мозга – аминокислоты. К возбуждающим относятся глутамат и аспартат. При освобождении в синапс они через ионотропные рецепторы открывают быстрые натриевые каналы, что приводит к быстрому входу в постсинаптический нейрон ионов натрия. Это деполяризует плазматическую мембрану и вызывает возбуждение нейрона. Возбуждающие аминокислоты необходимы для всех функций мозга, включая поддержание его тонуса, бодрствования, психологической и физической активности, регуляцию поведения, обучение, память, восприятие чувствительных и болевых импульсов. Однако их избыток вызывает тяжёлые заболевания. Например, избыток глутамата приводит к судорожным приступам. При ишемии в синапс выделяется так много глутамата, что вызывает чрезмерное накопление ионов Ca2+ в постсинаптическом нейроне и его перевозбуждение (нейротоксическое действие) – возникает инсульт [1, 41].
Ещё один возбуждающий медиатор – ацетилхолин (АХ), активирующий ионотропные N-холинорецепторы с открытием тех же быстрых натриевых каналов. Через эти рецепторы АХ участвует в функциях базальных ганглиев головного мозга, связанных с регуляцией двигательной активности и мышечного тонуса.
Главный ингибирующий нейромедиатор головного мозга гамма-аминомаслянная кислота (ГАМК). Образуется из глутамата, связывается с ГАМК ионотропными ГАМКА рецепторами (субъединицами хлоридных каналов), что приводит к их открытию и быстрому входу в постсинаптический нейрон ионов Cl-. Эти ионы вызывают гиперполяризацию и в результате - торможение нейрона
Аминокислота глицин – особый ингибирующий нейромедиатор спинного мозга. Он действует по аналогичному механизму, а антагонистом его рецепторов является стрихнин. [36, 37, 38, 38, 40, 42, 43].
К нейромодуляторам относятся все нейромедиаторы, но их модулирующие эффекты реализуются не через ионно-, а через метаботропные рецепторы. Ацетилхолин через М-холинорецепторы включает 3 разные СТС, что снижает уровень цАМФ, открывает К+ - каналы и вызывает накопление липидных вторых посредников и затем ионов Са2+. Через М-холинорецепторы (их в мозге больше чем N-рецепторов) АХ стимулирует образование условных рефлексов и память. Так при болезни Альцгеймера характерна ранняя гибель холинергических нейронов. Через эти рецепторы АХ реализует активность мотонейронов спинного мозга и регуляцию внутренних органов парасимпатическими нервами системы. ГАМК и её синтетические агонисты через оба типа рецепторов ГАМКА и ГАМКВ – снижают активность головного мозга.
Существует большое количество специализированных нейромодуляторов. В головном мозге из прогестерона образуются активирующие мозг нейромодуляторы нейростероиды. В отличие от большинства стероидных гормонов они действуют не путём проникновения в ядро клетки и соединения с ядерными рецепторами, а в результате активации ГАМКА– рецепторов нейронов [38, 41, 43].
Описанные 3 типа СТС опосредуют действие и некоторых других ингибирующих модуляторов, в том числе и пока единственного нуклеозидного НТ – аденозина. Через свои А1-рецепторы он снижает концентрацию ионов Са2+ в нейронах, что ингибирует высвобождение многих НТ, снижает тонус головного мозга, способствует утренней вялости [43, 44, 45].
Важный класс нейромодуляторов – моноамины: катехоламины (КА) и индолилалкиламины. КА синтезируются из аминокислоты тирозина, активность ключевого фермента синтеза тирозингидроксилазы увеличивается системой цАМФ протеинкиназа А. КА обеспечивают функционирование симпатико-адреналовой системы. Дофамин освобождается в основном в синапсах базальных ядер головного мозга, норадреналин – в стволе мозга и окончаниях симпатических нервов. Дофамин тормозной модулятор, снижающий эффекты возбуждающего медиатора АХ [38].
Норадреналин вызывает накопление в клетке ионов Са2+ (через α1-адренорецепторы) и цАМФ (через β-адренорецепторы). Активируется ретикулярная формация ствола, что тонизирует головной мозг, включая кору больших полушарий.
Однако те же КА через α2-адренорецепторы снижают концентрацию ионов Са2+ и цАМФ, что приводит к уменьшению выделения норадреналина и других НТ. ГАМК, аденозин и селективные антагонисты α2-адренорецепторов реализуют и другую приспособительную стратегию – толерантную. Для не характерно снижение потребления кислорода, температуры тела и катаболизма с уменьшением активности головного мозга и других физиологических систем. В результате значительно увеличивается устойчивость организма ко многим экстремальным факторам.
Индолалкиламины образуются из аминокислоты триптофана: серотонин в стволе головного мозга и энтерохромаффинных клетках кишечника, мелатонин – в эпифизе. Серотонин – снижает агрессивность, страх, депрессию, стимулирует пищевое поведение, сон и снижает болевые условные рефлекса, способствует обучению и лидерству. Мелатонин преимущественно вырабатывается ночью и способствует сну, тормозит выделение гонадотропных гормонов. Оба индолалкиламина снижают половую активность [39].
Наряду с этим большие успехи, достигнутые в последнее десятилетие в изучении синаптических молекул позволяют выделить группу атипичных трансмиттеров или нейрональных мессенжеров. К таким соединениям относят газ оксид азота (NO), цинк, выполняющий функции антагонистического котрансмиттера NMDA-рецепторов, а также локализованный в ганглиальных клетках d-серин, являющийся эндогенным лигандом места связывания глицина, входящего в состав NMDA-рецепторного комплекса [36, 37, 38, 38, 40, 44, 45, 46].
Метаболическая, миогенная и гуморальная регуляция
Миогенная регуляция мозгового кровотока осуществляется за счёт изменения внутриартериального давления и прямой ответной реакции мышц сосудистой стенки в виде сужения артерий при его повышении или их расширения при его снижении (феномен Остроумова - Бейлисса). Действие этой реакции кратковременно [26, 36].
Важной особенностью головного мозга является его высокая метаболическая активность. При поражениях головного мозга нарушения кровообращения приводят к сопряженным изменениям метаболических процессов независимо от характера повреждающего воздействия - травмы, компрессии, гипоксии мозга. Нарушения кровообращения и метаболизма мозга развиваются в определенной последовательности - накопление лактата, тканевой ацидоз, паралич вазомоторов, нарушение ауторегуляции, нарушение регуляции внутричерепного давления, исчезновение реакции церебральных сосудов на изменение концентрации углекислого газа, утрата контроля метаболизма, развитие отека мозга.
Активным фактором, влияющим на сосудистую стенку, является напряжение углекислоты в артериальной крови. Механизм действия СО2 на сосуды мозга изучен в работах многих авторов [27, 32, 47].
Величина просвета артерий зависит от напряжения СО2 в капиллярах и тканях, концентрации ионов Н+ в околососудистом пространстве и напряжения О2. Повышение напряжения СО2 вызывает выраженную дилатацию сосудов. Так, при повышении рСО2 вдвое мозговой кровоток также удваивается. Действие СО2 опосредовано соответствующим увеличением концентрации Н+, образующихся при диссоциации угольной кислоты. Прочие вещества, при накоплении которых увеличивается концентрация ионов водорода, также усиливают мозговой кровоток. Уменьшение напряжения кислорода вызывает расширение сосудов, а увеличение - сужение. Однако влияние рО2 на просвет сосудов ниже, чем влияние рСО2. Влияние углекислоты на церебральную гемодинамику объясняется либо непосредственным влиянием на сосудистую сеть, расположенную вблизи артериального конца капилляров [44, 48, 49], либо изменениями рН цереброспинальной жидкости, окружающей артериолу.
Многие авторы полагают, что действие СО2 на сосуды мозга реализуется с помощью нервных механизмов [29, 45, 46]. Существенная роль в механизме действия СО2 отводится также местным нервным механизмам [21, 24, 47]. Реакция мозгового кровотока на гиперкапнию ослабевает при гипотензии, ишемии мозга [48, 49, 50, 51, 52]. Этот эффект объясняют дилатацией артерий под действием накопившихся в условиях патологии кислых продуктов метаболизма и уменьшением резервных возможностей для их дальнейшего расширения [53, 54].
Влияние кислорода на цереброспинальное кровоснабжение выражено слабее по сравнению с СО2. Однако гипероксия и избыточная оксигенация крови ведут к сужению сосудов мозга, а гипоксия наоборот вызывает дилатацию и увеличение церебрального кровотока [55, 11, 44, 18]. Считают, что вазоконстрикторная реакция при гипербарической оксигенации носит приспособительный характер, защищая мозг от проникновения в него избыточного кислорода [48, 56].
В процессе адаптации сосудистого тонуса мозга к гипоксии предполагается участие тканевого метаболизма путем увеличения анаэробной передачи продукции АТФ в гладких мышцах мозговых сосудов [53, 57].
В литературе имеются данные об участии ионов К+ и Н+ в регуляции тонуса сосудов мозга [58, 44, 53, 59, 60]. Биохимические исследования [44] свидетельствуют о том, что под влиянием Са2+ интенсивность потребления кислорода и величина дыхательного контроля мозга повышаются, в тоже время увеличение концентрации ионизированного Са2+ в растворимой цитоплазме обеспечивает метаболические эффекты синаптической передачи нервных импульсов. Ионы Са2+ являются важным компонентом сосудистой стенки, запускающим сократительный мышечный механизм. Концентрация Са2+ оказывает влияние на диаметр пиальных артерий: увеличение ее приводит к их сужению, а снижение к расширению этих сосудов [61].
К метаболическим факторам регуляции мозгового кровотока относятся сдвиги газового состава крови, напряжения в ней кислорода, углекислоты [25, 62, 63]. Изменения газового состава крови изменяет рН среды, окружающей сосуды [64], вызывает сдвиг концентрации различных вазоактивных веществ [65, 66].
При различных изменениях системной гемодинамики роль данного механизма уникальна.
Монооксид азота образуется в эндотелии сосудов, способствует расширению микрососудов и является постоянным регулятором микроциркуляции. Согласно данным [67], большое количество этого вещества образуется в эндотелии капилляров. Угнетение его синтеза существенно изменяет скорость кровотока в капиллярах мышц. Предполагается, что угнетение синтеза монооксида азота в эндотелии после тяжелой ишемии ведет к массовой адгезии нейтрофилов к стенкам микрососудов и к ухудшению или прекращению их «проходимости» для крови.
В регуляции МК проявляют себя гуморальные механизмы, которые разделяют на 2 группы: 1 – гормоны, вырабатываемые нервными окончаниями железистых и тучных клеток, действие которых направлено на тонус прилежащих сосудов; 2 – гормоны, вырабатываемые специализированными органами внутренней секреции [68]. Гуморальные факторы обладают выраженными вазоактивными свойствами. Брадикинин вызывает у человека церебральную дилатацию и увеличение кровотока [69].
Под влиянием ацетилхолина в опытах описано как расширение, так и сужение пиальных артерий [22, 70, 71, 72, 73]. Аппликация серотонина на поверхность мозга вызывает длительную констрикцию крупных пиальных артерий [11, 23, 56, 74] за счет преимущественного действия на Д-рецепторы сосудистой стенки. Отмечено двухфазное действие дофамина – первичное сужение с последующей дилатацией [11, 75, 76, 77]. Важная роль принадлежит b-адренорецепторам в регуляции тонуса сосудов и метаболизма мозга у человека [73, 78]. Установлены региональные различия в распределении a- и b-адренорецепторов в сосудах мозга. Каротидная артериальная система более чувствительна к норадреналину, чем позвоночная [11, 21]. Простагландины вызывают неоднозначные реакции на внутри- и внечерепные сосуды. Теория, объясняющая многосторонние действия простагландинов основаны на способности их влиять на синтез циклических нуклеотидов, главным образом цАМФ [11, 79].
Метаболизм головного мозга носит аэробный характер и для обеспечения жизнедеятельности и анатомической целостности его структуры мозгу необходимо до 20% всей циркулирующей крови и 25% всего кислорода. Этим определяется такой высокий кровоток, высокий уровень потребления кислорода, многоуровневая, дублированная система регуляции мозгового кровообращения и механизмы компенсации при поражении одного или нескольких бассейнов кровоснабжения. При полном прекращении поступления кислорода и глюкозы, вследствие нарушения кровотока или при других причинах через 1-3 секунды происходит потеря сознания, через 4-6 минут необратимая гибель мозга. Период 4-6 минут критический. При этом последующая перфузия не приводит к восстановлению кровотока на различных территориях мозга вследствие перекрытия капиллярного отдела микроциркуляторного русла. Перемещение крови происходит из областей мозга, менее активных в функциональном отношении, в области с интенсивной деятельностью. Величина локального кровотока в это время значительно повышается в одних областях, снижаясь одновременно в других на фоне стабильного или, реже, несколько увеличенного кровотока в мозге в целом. Но этот критический период в 4-6 минут может расширяться до десятков минут при переохлаждении и наличии в организме седативных веществ (барбитураты, нейролептики, транквилизаторы). Следует отметить, что при падении парциального давления кислорода ниже 65 мм рт.ст. нарушается синтез медиаторов и начинаются изменения сознания [80]
1.2 Роль селена в организме
Селен является эссенциальным микроэлементом для человека и животных. Он является одним из биологически важных микроэлементов, присутствующих в организме человека и участвующих в метаболических, биофизических и энергетических реакциях организма, обеспечивающих жизнеспособность и функции клеток, тканей, органов и организма в целом. Особенно важна роль селена для функциональной активности таких органов как сердце, печень, почки и др.
Селен – элемент 4 группы главной подгруппы периодической системы Менделеева, во многом повторяющий химические свойства серы. Селен способен замещать серу в серосодержащих аминокислотах с образованием селеноаминокислот, которые активнее в биологическом отношении, и являются более сильными протекторами ионизирующей радиации, чем серосодержащие аминокислоты. Кроме того, селеноаминокислоты способствуют уменьшению количества свободных радикалов, нарушающих активность и свойства ферментов и аминокислот [81, 82, 83].
Селен поступает в организм человека из почвы с продуктами растениеводства и животноводства, что определяет зависимость уровня обеспеченности микроэлементом от геохимических условий проживания.
Однако не весь селен почвы доступен для растений. Так, в кислых, сильно заболоченных почвах биодоступность микроэлемента низка, хотя общее содержание может быть и значительным [83].
С учётом того, что оптимальный уровень потребления селена, соответствующий максимальной активности глютатионпероксидазы (GPX) тромбоцитов или содержанию селена в сыворотке крови 115-120 мкг/л, составляет 120 мкг/сут, установленные концентрации селена соответствуют умеренной обеспеченности населения микроэлементом на большинстве исследованных территорий, причём ни в одном из регионов не зарегистрированы случаи глубокого дефицита селена – содержание в сыворотке крови менее 50 мкг/л. В России средние концентрации селена в сыворотке составляют от 62 мкг/л на западе, до 145 мкг/л на востоке [81, 84, 85].
У растений важнейшей химической формой селена является селенометионин. Большая часть селена в животных тканях присутствует в виде селенометионина и селеноцистеина [86].
Биохимические функции селена определяют селенсодержащие белки (СБ). Недостаток микроэлемента может приводить к нарушению клеточной целостности, изменению метаболизма тиреоидных гормонов, активности биотрансформирующих ферментов, усилению токсического действия тяжёлых металлов, повышению концентрации глютатиона в плазме [87, 88].
Характерной особенностью СБ млекопитающих является то, что они, по-видимому, связаны с окислительно-восстановительными процессами, проходящими внутри клетки и вне ее [89]. К настоящему времени охарактеризованы 12 СБ, содержащих в активном центре селен [90].
- GPX1 (cCPX) клеточная глутатионпероксидаза – предполагается её наличие во всех клетках организма млекопитающих, по-видимому используется как селеновое депо, антиоксидант.
- GPX2 (CPX-СI) – локализуется в клетках эпителия желудка
- GPX3 (рCPX) межклеточная GPX или GPX плазмы, контролирует уровень перекисей вне клетки, функция фермента не выяснена, однако показано, что активность рCPX восстанавливается быстрее, чем cCPX, что может говорить о большей значимости этого фермента.
- GPX4 (РНCPX) фосфолипид, локализуется в основном в семенниках, однако найден в мембранах, цитозоле. Восстанавливает гидроперикиси холестерина, его эфиров, фосфолипидов, играет важную роль в репродуктивной системе мужчины [91].
- ID – группа 3 оксидоредуктаз, регулируют активность тироксина. В экспериментах на животных показано, что одновременный дефицит селена и йода приводит к более сильному гипотиреоидизму, чем дефицит одного йода. Некоторые авторы предполагают, что кретинизм у новорожденных может быть следствием комбинированного дефицита этих 2 элементов у матери [87].
- ID1 – фермент, участвует в метаболизме тироксина и трийодтиронина. Это микросомальный фермент локализован в печени, почках, щитовидной железе и ЦНС.
- ID2 – катализирует превращение тироксина в трийодтиронин
- ID3 – дезактивурует тироксин и трийодтиронин, локализован в ЦНС, коже, плаценте. Участвует в метаболизме энергии [92].
- TR млекопитающих – основная функция – катализирует NADPH – зависимое восстановление в цитозоле.
- SPS2 – фермент, катализирует АТФ-зависимую активацию селена с образованием селенофосфата.
- SelP – гликопротеин, может выполнять роль антиоксиданта и селенового депо. Быстро синтезируется при введении селеновых добавок. Участвует в дезактивации тяжёлых металлов.
- Селенопротеин W (SelW) межклеточный белок, присутствует во многих тканях преимущественно в мышцах и мозге. Предполагается его участие в окислительно-востановительных реакциях, влияние на развитие онкологических заболеваний.
Данные изотопного анализа и результаты теоретических исследований позволяют предполагать, что в организме млекопитающих может насчитываться от 20 до 100 СБ [93].
Повышение заболеваемости раком и сердечно-сосудистыми заболеваниями при дефиците селена, бесплодие у мужчин и увеличение риска смерти от СПИДа могут быть связаны со снижением биосинтеза СБ и нарушением соответствующих биохимических процессов [81].
Согласно современным представлениям, общей регулируемой формой селена в организме является селенид, который образуется из селеноцистеина под действием Sec-β-лиазы [94, 95]. Предшественником селеноцистеина может являться селенометионин. Неорганический селен (селенит) реагирует с восстановленной формой глутатиона (GSH) также с образованием селенида [96]. Последний частично включается в биосинтез СБ и тРНК в результате реакции с селенфосфатсинтетазой (SPS), частично экскретируется из организма преимущественно в виде метилированных форм с мочой и дыханием [97, 98]. Фосфорилирование селенида осуществляется с участием АТФ [99]. Регулирование реакции фосфорилирования селенида определяет возможность депонировать селен – явление, наблюдаемое при дефиците микроэлемента. Ингибирование реакции приводит к увеличению концентрации селенида и как следствие, к увеличению экскреции селена. Эта ситуация наблюдается, когда селен доступен в количествах больших, чем необходимо для синтеза селенопротеинов [100, 101].
Абсорбирование селена организмом происходит в тонкой кишке, среди сегментов которой несколько большую скорость транспорта обеспечивает двенадцатиперстная кишка, откуда низкомолекулярные формы селена способны перейти в кровь уже через 1 минуту после поступления в кишку [102]. Абсорбция селенита натрия происходит отлично от органических соединений. Экспериментальные данные указывают на то, что селен вступает в неферментативную реакцию с GSH с образованием селенидиглутатиона [103], который может служить субстратом для γ-глутамилтрансферазы и таким образом переносится через мембраны клеток. Поскольку селеновый статус экспериментальных животных почти не влияет на величину абсорбирования вводимого селенита [103], следует предположить, что для этого соединения регуляторный механизм абсорбции отсутствует. Количество и распределение СБ в органах и тканях млекопитающих зависит от специфичности их экспрессии, селенового статуса организма, длительности приёма селена и химической формы селена в рационе [104].
При дефиците селена уровень СБ снижен, однако включение микроэлемента осуществляется в первую очередь в наиболее важные белки и ткани репродуктивные и эндокринные органы, мозг. Скелетные мышцы и сердце снабжаются селеном медленнее [99]
M. Wenzel и соавт. (1971) определили биологические полупериоды существования селена в тканях. В частности, для мышц этот срок составил 100 сут, для печени – 50 сут, почек – 32 сут и для сыворотки крови – 28 сут.
В условиях выхода из селендефицитного состояния активность GPX-GI достигает максимума уже через 10 часов после начала введения селена, тогда как активность cGPX начинает возрастать только через 24 часа и не достигает максимума даже через 3 дня [105].
Гомеостатическое регулирование уровня селена в различных органах и тканях приводит к тому, что при введении высоких доз селена уровень СБ превышает достигаемый при адекватном потреблении. У человека активность pGPX достигает максимума при потреблении всего 50 мкг селена в сутки [106].
При введении селенита натрия животным в высоких дозах не наблюдали увеличения активности фермента, несмотря на значительное возрастание концентрации микроэлемента в плазме и эритроцитах, но отмечали даже некоторое его снижение [107].
При снижении общего содержания селена в плазме и эритроцитах увеличивается доля PHGPX, а в эритроцитах возрастает уровень сGРХ и гемоглобина [108].
После введения радиоактивного селена значительная его часть связывается белками плазмы крови. При этом оказалось, что эритроцитам в данном процессе принадлежит ведущая роль, так как 75Se в виде селенита чрезвычайно быстро, в пределах нескольких секунд проникает через их мембраны [McMurray C.H., Davidson W., 1974]. Уже через 1-2 минуты в эритроцитах концентрируется 50-70% всего селена крови. На модели in vitro показана временная зависимость перераспределения селена между элементами крови. Есть основания полагать, что к 4 минуте концентрация микроэлемента достигает максимума. Затем в течение 15-20 мин почти весь селен выходит из эритроцитов, связываясь сначала с альбуминами, а затем с глобулинами плазмы крови [81, 109].
В эритроцитах присутствует селеновый «насос» у человека и ряда животных [Wright P.L., Bell M.C., 1963; K.J., Hidiroglou M. 1972; Sandholm., 1975]. Под влиянием системы глутатион – глутатионпероксидаза селенит подвергается превращению с образованием комплекса селена с глутатионом. При последущем восстановлении селен катализирует транспорт электронов к кислороду. Выйдя из эритроцита, возможно, в составе селеноглутатионового комплекса, этот микроэлемент фиксируется в белках плазмы. Кроме того, сниженная активность глутатионпероксидазы в эритроцитах, по-видимому, способствует образованию окислительных форм белков, например гемоглобина (HbSSG). Дефицит селена может приводить к гемолизу эритроцитов [109].
У соединений селена выявлена различная биодоступность. Установлено, что селен, содержащийся в большинстве исследованных соединений обладает меньшей биодоступностью по сравнению с селенитом натрия [110].
Селен выводится из организма в основном с мочой, фекалиями и выдыхаемым воздухом (чесночный запах). Среди путей выведения доминирующим является первый, а последний характерен при остром и хроническом отравлении. При токсикозах альтернативным путём выведения селена можно считать его накопление в волосах и ногтях [100].
Концентрация селена в моче в течение суток значительно меняется, однако большая часть вводимого селена экскретируется в течение 24 часов [101], что позволяет использовать этот показатель в качестве критерия обеспеченности селеном, т.к. он хорошо коррелирует с уровнем потребления этого микроэлемента. Обычно этим путём выводится около 40-50% потребляемого селена, однако в некоторых случаях эта величина может достигать 60%. В зависимости от потребляемой дозы концентрация селена в моче может варьировать от 0,9 мкг/л (эндемические зоны Китая) до 3900 мкг/кг (Венесуэла) [111].
Фактором, влияющим на уровень выведения, является химическая форма селена. Обычно неорганические соли легче выводятся из организма, что делает их более безопасными при потреблении, чем органические соединения. Есть данные, свидетельствующие о низком уровне выведения органических форм селена и, следовательно, о наибольшей опасности отравления при потреблении аномально высоких доз [112].
У здоровых добровольцев в нагрузочном тесте при ежедневном двукратном увеличении уровня потребления микроэлемента приём селенита натрия в дозах 100 – 800 мкг/сут. приводит к активной экскреции избытка селена с мочой, достигающей 80-90% от величины потребления.
При приёме препаратов органического происхождения предел выведения селена с мочой достигается при дозе 400 мкг/кг [81, 113].
Дефицит селена вызывает ряд эндемических заболеваний у человека и животных. «Беломышечная» болезнь (алиментарная мышечная дистрофия) характеризуется очаговой дегенерацией различной степени тяжести и некрозом скелетной и сердечной мышц невоспалительного характера, она предупреждается включением в рацион селена [81]. Патоморфологические изменения при этом заболевании характеризуются глубокими нарушениями скелетных мышц и миокарда. В частности, наблюдается пёстрая патогистологическая картина за счёт неравномерного полнокровия, дистрофических и некробиотических изменений кардиомиоцитов, нередко с явлениями дистрофического обызвествления. По мнению А.П. Авцына (1972), белая окраска мышц обусловлена исчезновением миоглобина и вторичным коагуляционным некрозом миоцитов. Изменения миокарда и скелетных мышц имеют дегенеративно-некробиотический характер. Болезнь «Кешана» представляет собой эндемическую фатальную кардиомиопатию, для которой характерны аритмии, увеличение размеров сердца, фокальные некрозы миокарда, за которыми следует сердечная недостаточность. У больных, страдающих данной болезнью выявляются аномалии мембран эритроцитов. В эритроцитах больных детей уровень селена, активность Na+, К+-АТФазы, текучесть липидов и их мембран отличаются от показателей детей контрольной группы, проживающих в том же регионе [114, 115, 116, 117, 118, 119].
При проведении в Финляндии в течение 5 лет эпидемиологических исследований на 11000 мужчин и женщин в возрасте 35-59 лет было установлено, что за этот период 252 перенесли инфаркт миокарда и 131 умерло от сердечно-сосудистых заболеваний. Во всех случаях уровень селена составил 52 мкг/л, в контроле 55 мкг/л. Ряд работ проведённых в ещё в 80-х годах продемонстрировал, что при концентрации селена в сыворотке ниже 0,4 мкмоль/л вероятность возникновения инфаркта миокарда увеличивается в 7 раз, а при содержании 0,4-0,6 мкмоль/л – в 3 раза [120].
В другом исследовании, проведённом в тех же условиях, для группы умерших уровень селена составил 62 мкг/л. В контроле 68 мкг/л. Относительный риск смерти при концентрации селена в плазме менее 45 мкг/л составил 3.2 [121].
В районах Центральной Африки, дефицитных одновременно по селену и йоду зарегистрирован эндемический микседематоидный кретинизм [122].
Экспериментальные и клинические исследования показали, что этиология кистозного фиброза поджелудочной железы (муковисцидоз) обусловлена дефицитом ряда элементов, особенно селена, в перинатальном периоде. Это заболевание распространенно среди детей младшего возраста. Кроме того, при дефиците селена наблюдается алиментарный гепатоз - некротические изменения печени, обширный отёк и отложение цероидного пигмента в жировой ткани и очаговая и диффузная инфильтрация в кишечнике, желудке, брыжейке и регионарных лимфатических узлах - идиопатическая эозинофильная инфильтрация [81, 123].
Первые сведения о селене связаны с проявлениями его токсичности, обусловленной аномально высоким потреблением. Выделяют несколько степеней токсичности [124, 125].
Острая токсичность проявляется при кратковременном потреблении высоких доз селена и быстро приводит к смерти. Признаки: чесночное дыхание, летаргия, избыточное выделение слюны, дрожание мышц, миокардит и т.д.
Подострая токсичность связана с потреблением высоких доз селена в течение значительного времени. Признаки: слепота, атаксия, дезориентация, затруднение дыхания [125].
Хронический селеноз развивается при потреблении умеренно-повышенного количества селена в течение нескольких недель или месяцев [114, 127].
Оценка степени токсичности соединений селена для человека затрудняется отсутствием селективного и чувствительного индикатора избыточного поступления селена в организм человека. Одним из возможных показателей является алопеция и изменение ногтей, а также преимущественное накопление селена эритроцитами по сравнению с плазмой [115].
Безопасный и достаточный уровень суточного потребления селена составляет 50 – 200 мкг/сут. Минимальная потребность в селене установлена по данным для эндемических регионов Китая: наименьшая величина потребления микроэлемента, при которой не наблюдалось развитие болезни Кешана, составила 19 и 14 мкг/сут для мужчин и женщин соответственно [117].
Физиологическая потребность в селене установлена по показателю потребления, обеспечивающему максимальную активность GPX плазмы. Для жителей биогеохимических провинций Китая с глубоким дефицитом селена эта величина составляет 40 мкг/сут [117]. Для европейцев этот уровень составляет 70 мкг для мужчин и 55 мкг для женщин [127, 138].
В Финляндии с учётом многолетнего опыта использования обогащённых селеном удобрений предполагается значительно более высокий уровень потребления селена, отвечающий физиологической потребности, а именно 120 мкг/сут, этому значению соответствует максимальная активность GPX тромбоцитов [85].
При расчёте РД (референт дозы) исходя из данных полученных при изучении эндемического селеноза в Китае берут 853 мкг/сут при массе тела 55 кг. Введение дополнительного коэффициента (х3) для учёта индивидуальной чувствительности даёт значение 5 мкг селена на 1 кг массы тела в сутки, что соответствует 350 мкг/сут при массе тела 70 кг [85, 111, 116, 128].
1.3 Фармакологическая характеристика селенсодержащих соединений
Биологические функции селена огромны. Антиоксидантные свойства селена определяют перспективность использования препаратов микроэлемента при оксидантном стрессе. Окислительное повреждение тканей играет ключевую роль в развитии многих заболеваний: атеросклероза, ишемической болезни сердца, диабетических ангиопатий, нейродегенеративных и аутоиммунных заболеваний, рака, лучевой болезни [129, 130, 131, 132], псориаза [134], ожогов [135], катаракты [136, 137] и др. Для фармакологической коррекции окислительного стресса широко используются антиоксиданты различной химической природы. По мнению В.Г Зайцева, О.В. Островского, В.И. Закревского наибольшие перспективы клинического применения имеют представители групп катализаторов и ловушек радикалов. Антиоксиданты – катализаторы не расходуются в ходе защитных реакций, а значит, могут быть использованы в существенно меньших дозах, чем АО других групп. Их эффект в организме будет сохраняться более длительное время, а вероятность побочного действия у них гораздо меньше. Кроме того, пока нет данных о возможности проявления АО данной группы прооксидантного действия в условиях близких к физиологическим. Наибольшие перспективы в медицинском применении имеют имитаторы глутатионпероксидазы, к которым относят селенсодержащие соединения. Известно, что ГП катализирует превращение опасных для организмов гидропероксидов (ROOH) и Н2О2 в инертные гидроксисоединения (RОН) и воду соответственно при участии глутатиона. Глутатионпероксидаза – первый селеносодержащий фермент, найденный в организме млекопитающих. Фермент не обладает строгой специфичностью по отношению к перекисям и нуждается в качестве кофактора в глутатионе, который в ходе ферментативной реакции подвергается окислению:
Н2О2 + 2GSH → H2O + GSSG
ROOH + 2GSH → ROH + H2O + GSSG
В этой реакции электрон переносится на перекись не с восстановленного глутатиона, а с селенола, который при этом переходит в селенистую кислоту, а затем последняя регенерирует в селенол восстановленным глутатионом. Имитаторы ГП эффективны в основном для снижения интенсивности ПОЛ [81].
Изучение терапевтического влияния селена на течение экспериментального инфаркта миокарда у крыс позволило чётко установить его благотворное действие [Кактурский Л.В., 1986]. Заживление инфаркта миокарда продемонстрировано в эксперименте на крысах, у которых инфаркт воспроизводили лигированием венечной артерии (120 животных). В течение недели после инфаркта крысам подкожно вводили водный раствор селенита натрия из расчёта 30 мкг/кг в сутки. Для потенцирования эффекта селена одновременно внутримышечно вводили α-токоферол в дозе 50 мг/кг в сутки. Крыс забивали через 2 ч, 1, 3, 7, 14, 30 суток. У леченых крыс отмечено уменьшение зоны инфаркта миокарда, особенно выраженное в ишемической стадии (4,52% против 31,7% в контроле, р<0,001). Величина зоны инфаркта коррелировала с уровнем перекисного окисления липидов в этой же зоне, определявшимся хемолюминисцентным методом. Гистостереометрически показано ускорение созревания гранулоцитарной ткани у леченых животных. Это выражалось увеличением числа клеток фибропластического ряда и уменьшением доли сегментоядерных лейкоцитов. Во внешней зоне инфаркта в отличие от контроля под влиянием селена отмечены нормальная активность оксидоредуктаз, а также сохранность ультраструктурной организации кардиомиоцитов, т.е. отмечен протективный эффект селена на кардиомиоциты на гисто- и ультраструктурном уровне. Сформированная на 30-е сутки постинфарктная зона у нелеченых животных имела рыхлое строение, а у крыс, получавших селен, - компактное строение со значительным содержанием мышечных элементов.
Из приведённых данных видно, что незначительное понижение селенового статуса ведёт к тяжелым нарушениям сердечно-сосудистой системы. В экспериментах показано, что процент постишемического восстановления механической функции сердца в группе с добавлением селена выше, а показатели дегенерации ткани вследствие ишемии ниже по сравнению с контролем.
30-летние исследования А.Н. Кудрина и соавт. убедительно доказывают, что селенит натрия можно рассматривать в качестве регуляторного элемента жизнедеятельности клеток. Авторами показана выраженная эффективность селенита натрия при инфаркте миокарда. Наибольшее торможение ПОЛ и протекторное действие на мембраны кардиомиоцитов обнаружила комбинация селенита натрия и α-токоферола вследствие потенцирования эффектов. Под влиянием этих ингибиторов ПОЛ динамика выздоровления ускорялась при уменьшенных размерах рубца за счёт ограничения поражения миокарда в околоинфарктной зоне. Кроме того Кудриным и соавторами получены данные согласно которым селенит натрия и органические соединения селена способны устранять различные формы аритмий и смертельную фибрилляцию сердца, вызаваемую хлоридом кальция и гистамином. Авторы делают заключение, что селенит натрия, α-токоферол, убихинон и особенно их комбинации являются главной антиоксидантной системой организма, защищающей его от мембранной патологии при ишемии, гипоксии, ионизирующем излучении, интоксикации ССl4, дистрофии [4, 138, 139].
По данным С.М. Николаева селенит натрия предохраняет миокардиальные клетки от разрушения, ограничивает периинфарктную зону, уменьшает размеры рубцов [3]. М.Д. Савиной и А.Н. Кудриным выявлен антиаритмический эффект селенита натрия на экспериментальных моделях аритмий [4]. Т.А. Венцславская и д.р. изучали эффекты препарата пипередин-этил-селенофена при экспериментальной аритмии сердца. Антиаритмическая активность сравнивалась с изотином (верапамил). Выявлено противоаритмическое действие вещества. Было установлено, что изоптин и пипередин-этил-селенофен, применённые в дозе 1 мг/кг для лечения нарушений ритма сердца, вызванных кальция хлоридом в дозе 125 мг/кг, вызывают во всех случаях лечебный эффект. При этом пиперидин-этил-селенофен восстанавливает ритм и его регуляторные механизмы значительно быстрее, чем изоптин. Выявлено кардиопротекторное действие селена на модели хронической нагрузки железом у мышей [140].
В эпидемиологических исследованиях отмечена обратная корреляция между уровнем селена в плазме и риском развития коронарной болезни сердца и атеросклероза. Снижение уровня селена в плазме коррелировало с увеличением свертываемости крови и повышением синтеза предшественников агрегации – эйкозаноидов, таких как тромбоксан А2 и лейкотриены [141].
Селен оказывает влияние на биосинтез простагландинов. Отмечено значительное увеличение времени свертывания крови у людей, потреблявших ежедневно 700 мкг селена в виде селенита натрия в течение 6 недель [142]. Другой аспект влияния селена на метаболизм простагландинов – защита простагландиндегидрогеназы, ключевого фермента деградации простагландинов. При нормобарической гипероксии, активность этого фермента снижается, благодаря чему наблюдается сосудосуживающий эффект. Такая защита может быть осуществлена введением в диету крыс витамина Е в дозе 600 МЕ/кг (сохраняется около 50% активности простагландиндегидрогеназы ткани лёгких при кислородной экспозиции) или селена в дозе 100 мкг/кг массы тела [143].
Показано, что эндотелиальные клетки с высоким содержанием селена не ингибируют агрегацию тромбоцитов в присутствии ацетилсалициловой кислоты [144].
Б.И. Левшин выявил нормализующее влияние селенита натрия, селенофена 5, селенофена 6, на изофермент ЛДГ5 и общую активность ЛДГ сыворотки крови. Препараты селена оказывают положительное влияние на показатели белкового, жирового и углеводного обмена при токсическом гепатите. Лечебно-профилактическое введение селенита натрия способствует некоторому ускорению регенерационных процессов в печени после её прижизненной частичной экстирпации, о чём свидетельствует более быстрое нарастание и увеличение содержания гликогена в печени [139, 145, 146, 147]. К.О. Шарипов выявил влияние органических производных селена в регуляции антиокислительных процессов в печени при экспериментальном токсическом гепатите [148].
Установлено, что при острых инфекционных заболеваниях уровень селена в сыворотке крови снижен. При этом отмечено, что максимальное снижение уровня селена отмечается у больных в тяжёлом клиническом состоянии [81, 149]. Показано, что одновременное обогащение рациона животных селенсодержащими дрожжами и витамином Е оказало стимулирующий эффект на В-систему иммунитета, выражающийся в увеличении количества антителобразующих клеток, и на митогенную активность Т-лимфоцитов. Иммуномодулирующий эффект соединений селена относят за счёт функции глутатионпероксидаз, которые обеспечивают восстановление гидроперекисей и других продуктов свободнорадикальных реакций и регулируют выход липоксигеназных и циклооксигеназных метаболитов арахидоновой кислоты [150, 151, 152].
Селен оказывает влияние на репродуктивную функцию. У всех изученных видов животных дефицит селена вызывает нарушение воспроизводительной функции. При дефиците селена самки крыс приносят всего 1-2 нежизнеспособных детёныша. Из всех органов яички содержат наибольшее количество селена. У сельскохозяйственных животных при применении добавок селенита натрия увеличивается рождаемость, улучшается выживаемость молодняка, снижается частота бесплодия. При дефиците селена течка, овуляция, оплодотворение и раннее развитие плода у овец протекают нормально, но на 3-4 неделе суягности эмбрионы погибают [153, 154, 155, 156].
Установлено, что половые гормоны, СТГ и эритропоэтин влияют на распределение селена в элементах крови. Процесс перераспределения селена под действием половых гормонов сопровождается изменениями иммунологических показателей крови [81, 156, 157, 158].
У селена описано радиопротекторное действие. Снижение уровня селена в сыворотке крови является характерной реакцией организма на облучение. Дополнительное введение неорганических соединений селена, в частности селенита натрия, с водой животным, подвергнутым облучению, приводило к увеличению средней продолжительности жизни и в значительной степени уменьшало частоту возникновения радиационно-индуцированных опухолей по сравнению с таковой у облучённых животных, не получавших селен [129, 130, 131, 132].
Препарат «Селена», получаемый из дрожжей и содержащий селен 100 мкг/таб. используется в качестве радиозащитного средства. Он обладает способностью уменьшать стимулирующее действие CCl4 на перекисное окисление липидов, снижает достоверно перекисное окисление липидов в 1,5 раза, одновременно повышает уровень активности супероксиддисмутазы в 3 раза, статистически достоверно не изменяет уровень активности каталазы, повышает уровень селена в печени [159].
Роль селена при онкологических заболеваниях значительна. Противоопухолевая активность соединений селена пропорциональна их каталитической активности. Установлено, что в ряду различных соединении селена наибольшая активность наблюдается у селенотрисульфида глутатиона. Это соединение было запатентовано в качестве канцеростатического препарата в связи с его высокой цитотоксической активностью по отношению к клеткам карциномы лёгких, аденокарциномы ободочной кишки, меланомы, аденокарциномы молочной железы, глиомы, медуллобластомы, опухольтрансформированных фибробластов и кератиноцитов [160]. Вторым по активности соединением является селенит. Значительное количество работ посвящено изучению антимутагенности селенита натрия [161, 162]. Было сделано предположение, что высокие уровни селена в рационе животных могут стимулировать репарацию повреждённой ДНК, вызываемую канцерогеном [163, 164, 165].
Также есть данные об антиканцерогенном действии фармакологических доз селена. В сороковых годах 20 в была установлена его защитная роль в отношении химически индуцированных опухолевых клеток [166, 167], а в шестидесятых годах появились первые результаты, свидетельствующие об обратной связи между уровнем обеспеченности селеном населения и показателями смертности от онкологических заболеваний. Специфической особенностью онкологических заболеваний является накопление селена опухолью за счёт уменьшения его концентрации в мозге, сердце и мышцах. Расчёт коэффициента корреляции между величиной смертности и уровнем селена в сыворотке крови показал наличие обратной корреляции для лимфом, рака желудочно-кишечного тракта, лёгких, молочной железы, толстой и прямой кишки, печени. В большинстве исследований отмечалось, что высокие дозы селена снижают частоту развития опухолей в значительном числе случаев более чем на 35%. Клинические исследования в США показали, что у пожилых людей потребление селена снижает риск раковых заболеваний на 65% [168, 169, 170].
По расчётам ряда авторов, контингент людей, с низким содержанием селена (1,63 мкмоль/л) имеет в 2 раза больший риск заболеть раком, чем люди с высоким уровнем селена в организме (норма 1,72 мкмоль/л) [171, 171].
Показано, что селен стимулирует апоптоз - программируемая клеточная смерть. Это может предотвратить закрепление мутаций в последующих поколениях клеток. Этот эффект наблюдается при применении высоких доз селена и связан с хемозащитным действием микроэлемента [173, 174].
Противоопухолевое действие селена нельзя объяснить исключительно его участием в антиоксидантной системе GPX. Исключение предраковых генетически повреждённых клеток за счёт апоптоза представляется более эффективным в предотвращении рака, чем подавление пролиферации. Клинический эффект химических препаратов, большинство из которых ингибирует пролиферацию клеток, прекращается при отмене приёма лекарства. Агенты, индуцирующие апоптоз, такие как селен, могут обеспечивать более быструю защиту с меньшей токсичностью [81].
Известен ряд эффектов взаимодействия селена с витаминами и микроэлементами. Предполагают, что витамин С восстанавливает селенит до элементарного селена, а элементарная сера и селен легко соединяются, образуя сульфиды и селениды, содержащие два и более атомов серы или Se. Предполагается также, что Sec, посредством переноса электронов может соединяться с элементарным селеном с образованием селенсодержащих связей [81, 172].
Степень абсорбции селена зависит также в значительной степени от потребления β-каротина и, по-видимому, других жирорастворимых витаминов, аккумулирование которых организмом происходит в тонкой кишке [175].
Витамин Е является сильным антиоксидантом, однако для подавления перекисного окисления путём использования только витамина Е является недостаточным для ингибирования образования опухоли. В исследованиях было показано, что совместное применение витамина Е и селена более адекватно. Так при приёме в диете 2,5 мг/кг селена снижает общее число опухолей на 45% у животных, находящихся на адекватной по витамину Е диете и только на 24% при дефиците витамина Е. Высказываются предположения о том, что витамин Е и селен совместно обеспечивают антиоксидантную защиту по какому-то другому пути. Взаимосвязь между селеном и витамином Е так же объясняется их воздействием на различные этапы образования органических перекисей [81, 172].
Л.А. Кудрявцева (1964-1969) показала, что селен и витамин Е являются необходимыми компонентами обмена веществ у животных. Но витамин Е фармакологически менее активен, он только в больших дозах 100 мг/кг вызывает эффекты, сходные с таковыми при малых дозах селенита натрия 100мкг/кг. Это соотношение варьирует в отдельных случаях. Например, при экссудативном диатезе цыплят 1 молекула селена способна заменить 700-1000 молекул витамина Е. Также отмечено, что Se замедляет распад витаминов А и Е. А при недостатке селена снижается и общее содержание микроэлементов в организме. Селенит натрия в дозе, составляющей 1% от хронической токсической дозы (300-400 мкг/ 100г рациона), в 500 раз активнее витамина Е и в 250000 активнее α-цистеина при некротической дегенерации печени [176, 177].
Описано положительное действие селена в отношении ряда ксенобиотиков. Описано протективное действие в отношении нитратов. Так, у животных получавшие селен и нитраты в рационе отмечено увеличение концентрации селена в сыворотке крови при снижении концентрации метгемоглобина [177, 178, 179, 180, 181].
Выявлено защитное действие селена в отношении тяжёлых металлов. H2Se способен вступать в реакцию с металлами, образуя нерастворимые комплексы, понижающие биологическую доступность селена и металла. Это взаимодействие лежит в основе снижения токсичности металлов повышенными дозами селена [182].
Селен проявляет защитное действие в отношении органических и неорганических соединений ртути. Так селенит натрия предотвращает некроз почек и снижает смертность, связанные с воздействием хлорида ртути и метилртути [183, 184]. Селен обеспечивает защиту от токсического действия кадмия, он полностью снимает его тератогенный эффект [185, 186].
Описано взаимодействие селена с мышьяком, который является антагонистом селена [187, 188]. В связи с тем, что свинец, олово, теллур имеют сходные структуры, они обладают аффинностью по отношению к соединениям серы и взаимодействуют с селеном. Экспериментальные данные указывают на возможность конкурентного действия селена и меди [189].
Образование биологически недоступных соединений селена с металлами объясняется способность серебра, кадмия и др. вызывать у животных вторичную недостаточность селена и блокировать синтез глутатионпероксидазы даже при рационах, содержащих адекватное количество селена [81].
В настоящее время всё большее количество работ посвящено исследованию взаимосвязи различных элементов в организме. Показано, что медь, цинк, селен и молибден вовлечены в значительное количество биохимических процессов [190]. Отмечено, что содержание крыс на полусинтетическом аминокислотном рационе с низким содержанием селена сопровождалось резким понижением уровня других микроэлементов, например цинка в клетках панкреатических отростков, в паренхиме почек и в клетках сперматогенного эпителия [153, 191].
Известно, что интенсивная физическая нагрузка определяет ускорение метаболических процессов, приводя к значительному оксидантному стрессу. Это состояние может быть нивелировано добавлением селена.
Изучение влияния препаратов селена на выносливость организма при физических нагрузках позволило выявить существование температурного эффекта действия селена. Так, при высоких температурах у животных получавших селен, выносливость была ниже на 35-38% (р<0,05), чем при низких температурах увеличивалась почти на 200%. (р<0,05) по сравнению с контролем.
Явление селеновой активации работоспособности при низких температурах имеет особое значение в связи с известными данными о высоком уровне антиоксидантной защиты именно у полярных животных (северный медведь, тюлени, моржи и д.р.), в организме которых в пищевой метаболизм активно включены жиры, а также о повышенном метаболизме жиров у местного населения Севера (эскимосы, чукчи). Высокий уровень селена в сыворотке крови этих народов позволяет предполагать тесную взаимосвязь между указанными явлениями. Вероятно, что положительный эффект от приёма селена следует ожидать, когда организм с чисто углеводного обеспечения мышечных усилий переходит на более долговременные источники энергетического снабжения мускульных сокращений, т.е. в условиях переохлаждения и длительных мышечных перегрузок [81].
Таким образом, анализ литературных данных, посвящённый селенсодержащим соединениям, демонстрирует их разностороннем фармакологическом влияние на организм. Экспериментальные исследования и клинические наблюдения показали, что селениты благоприятно влияют на антиоксидантный статус организма, обладают кардиопротекторной, гепатопротекторной, радиопротекторной активностью, предотвращают или тормозят развитие опухолевых процессов, а также целого ряда заболеваний связанных с дефицитом селена. Однако действие селенитов на мозговой кровоток, динамику развития постишемических феноменов и психоневрологический статус остаётся неизученным.
Указанные обстоятельства и послужили основанием для сравнительного изучения влияния селенита натрия и селенита цинка на системную гемодинамику и мозговой кровоток в условиях нормы и при патологических состояниях.
ГЛАВА 2 МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
2.1 Изучение острой токсичности
Определение острой токсичности проводили на беспородных белых мышах обоего пола массой 20-25 г. Животные прошли карантин в течение 14 дней. Исследуемые вещества вводили внутрибрюшинно. Наблюдение за поведением и состоянием подопытных животных проводилось в течение 14 суток, при этом отмечали внешний вид, поведенческие реакции и количество погибших животных [192].
Классифицировали группу токсичности согласно К.К. Сидорову [193].
2.2 Регистрация мозгового кровотока методом водородного клиренса
Объемную скорость мозгового кровотока регистрировали методом водородного клиренса с помощью вживленного платинового электрода, расположенного на поверхности сагиттального синуса в области стока синусов.
Метод основан на скорости вымывания предварительно введенного водорода из мозговой ткани и позволяет определить объемную скорость мозгового кровотока. Положительными сторонами метода являются отсутствие травматизации сосудов мозга, стабильность показателей, индифферентность используемого газа.
На основе водородного клиренса регистрировали мозговой кровоток на наркотизированных крысах. Результаты оценивались по кривой изменения напряжения водорода на электроде полярографическим способом [10, 30, 59, 194, 195].
2.3 Регистрации параметров кардиогемодинамики с помощью компью-терной программы «Bioshell» на бодрствующих животных
Опыты проводились на белых крысах. Предварительно за 24 часа до начала эксперимента (хлоралгидрат 300 мг/кг, внутрибрюшинно) имплантировали полиэтиленовый катетер через правую сонную артерию [196]. Периферический конец катетера подкожно выводили на холку животного и фиксировали. Регистрацию данных производили с использованием одноразовых датчиков SP-01 (США) и компьютерной программы «Bioshell» в реальном масштабе времени на базе персональных компьютеров IBM AT 486 и Pentium-133 [197]. Анализировали следующие параметры кардиогемодинамики:
- артериальное давление
- частоту сердечных сокращений.
2.4 Методика оценки поведения животного в тесте «открытое поле»
Оценку влияния исследуемых соединений на ориентировочно исследовательскую и двигательную активность животных проводили в тесте «открытое поле». Предложенный Холлом метод "открытого поля" широко применяется в различных экспериментальных исследованиях, связанных с изучением поведения, психофармакологией.
Установка представляет собой круг диаметром 80 см, с бортиком 20 см, разделённый на 8 радиальных секторов и одним в центре. Тестируемое животное помещали в центральный сектор, спиной к экспериментатору. В течение 3 минут наблюдения регистрировали число пересечённых секторов (двигательная активность), вставание на задние лапы и число пересечённых центральных секторов (ориентировочно-исследовательская активность), количество стереотипных движений (груминг), эмоциональную активность (дефекация, уринация) [192, 198, 199, 200, 201].
2.5 Оценка координации движений
Координацию движений крыс оценивали в тесте вращающегося стержня. Крыс помещали на горизонтальный стержень диаметром 4 см, вращающийся со скоростью 3 об/мин. Неспособность животных удерживать равновесие на стержне в течение трёх минут рассматривали как проявление нарушения координации движений [192, 202].
2.6 Методика условного рефлекса пассивного избегания (УРПИ)
Данная модель предназначена для изучения процессов связанных с воспроизведением памятного следа. Оценку антиамнестического эффекта проводили по измерению латентного периода захода животного в тёмный отсек при воспроизведении навыка УРПИ. Выработку условной реакции пассивного избегания затемнённого отсека производили в экспериментальной камере, которая состояла из двух смежных отсеков, большого освещенного 60 на 60 см, и малого затемнённого (15х15см), снабжённого электродным полом.
Отсеки сообщались между собой с помощью четырёхугольного отверстия (8х8см). Крысу помещали на середину площадки светлого отсека, хвостом к тёмному отсеку и в течение трёх минут регистрировали латентный период первого захода в тёмный отсек. Животных, не заходивших в тёмную камеру, по истечение трёх минут из опыта исключали.
В тёмном отсеке животное получало через электрический пол однократное электроболевое раздражение электрическим током (40 в), состоящее из трёх импульсов длительностью 1 сек, следующих с интервалом 0,5 сек. Крыса считалась обученной, если в течение 30 сек после сеанса обучения она не заходила в тёмный отсек экспериментальной камеры. Тест на воспроизведение УРПИ осуществляли через 24 часа после обучения при повторном помещении животных в светлый отсек камеры и регистрации в течение трёх минут латентного периода первого захода в тёмный отсек, количества заходов и общего времени пребывания в тёмном отсеке. В модификации Буреш, Бурешова выработку условной реакции пассивного избегания затемнённого отсека также производили однократным электроболевым раздражением электрическим током (40 в), состоящим из 12 импульсов длительностью 1 сек, следующих с интервалом 5 сек. в течение минуты. Модификация Буреш, Бурешова позволяет судить о влияния исследуемых соединений на процесс фиксации обучения и консолидацию памяти (переход кратковременной памяти в долговременную) поскольку обучение проводили после ишемии, а также о степени неврологических нарушений, животных перенесших тотальную ишемию мозга, т.к. способ обучения не требует абсолютной выработки рефлекса избегания [192, 203].
2.7 Методы изучения противогипоксической активности
Для создания патологического фона, на котором изучалась эффективность исследуемых соединений, использовались различные модели циркуляторных гипоксий.
Нормобарическая гипоксия с гиперкапнией (т.н. «баночная гипоксия») является наиболее простым методом оценки антигипоксической активности. Животных одинакового веса (разброс не более 2 г на группу) помещали в герметически закрытые (крышки смазывали вазелином) банки 750 см3. Во всех случаях регистрировали время выживания (смерти) животных [202].
Циркуляторную гипоксию воспроизводили путём перевязки двух передних общих сонных артерий. Операция проводилась в асептических условиях с использованием наркоза хлоралгидрат, внутрибрюшинно, 300 мг/кг массы тела. За оперированными животными наблюдали в течение 3 суток с регистрацией числа выживших животных в опыте и контроле [192].
Следующую модель ишемии мозга создавали критическими гравитационными перегрузками в кранио-каудальном положении животных. Известно, что продольные гравитационные перегрузки, создаваемые центрифугой, вызывают нарушения кровоснабжения головного мозга, степень и характер которых зависит от величины и вектора ускорения.
Крысы помещались в отдельные продольные ячейки, объём ячеек соответствовал размеру животных и не давал возможности самопроизвольно отклоняться от заданного вектора ускорения.
Центрифуга представляет собой мотор с приводом установленный на станине. К приводу сверху крепится коромысло длиной 2,0 м, по краям которого расположены камеры для животных. Также имеется пульт управления, позволяющий контролировать число оборотов, градиент нарастания и спада перегрузок [204].
2.8 Методика воспроизведения постишемических цереброваскулярных феноменов
Для оценки действия исследуемых препаратов на динамику развития постишемических цереброваскулярных феноменов воспроизводили ишемию мозга путем двухсторонней окклюзии сонных артерий в течение 10-12 мин. при снижении САД до 40 мм рт.ст. После ишемии и реинфузии крови в артериальную систему животного осуществляли регистрацию скорости МК методом клиренса водорода [10, 30, 59, 194, 195].
2.9 Условия экспериментальных исследований
Экспериментальные исследования проведены на 236 крысах массой 200 -250 г обоего пола и 154 белых мышах массой 20-25г обоего пола. Исследовали неорганические производные селена: селенит натрия, селенит цинка.
В связи с тем, что селенит натрия и селенит цинка легкорастворим в воде в экспериментах использовали водные растворы. Растворы веществ готовили добавлением при перемешивании физиологического раствора (0,9% раствор натрия хлорида).
2.10 Статистическая обработка данных
В таблицах диссертации данные представлены в виде средних арифметических и ошибки среднеквадратичного отклонения. Исходные данные приведены в абсолютных значениях, а изменения, которые произошли после введения препаратов, приведены в процентном отношении к исходным показателям. Статистическую обработку полученных результатов производили по критерию Стьюдента [192]. Различия считались достоверными при уровне значимости р<0,05, р<0,01 для парных выборок по критерию Стьюдента.
ГЛАВА 3 ВЛИЯНИЕ СЕЛЕНИТА НАТРИЯ И СЕЛЕНИТА ЦИНКА НА ЦЕРЕБРАЛЬНУЮ ГЕМОДИНАМИКУ, СИСТЕМНОЕ АРТЕРИАЛЬНОЕ ДАВЛЕНИЕ И ЧАСТОТУ СЕРДЕЧНЫХ СОКРАЩЕНИЙ УСЛОВИЯХ ЭКСПЕРИМЕНТАЛЬНОЙ НОРМЫ
Острые опыты проведены на 90 мышах массой 20 – 22 г и 48 крысах массой 200 – 250 г. Регистрацию объёмной скорости мозгового кровотока осуществляли методом водородного клиренса. Системное артериальное давление и частоту сердечных сокращений у бодрствующих животных проводили с использованием одноразовых датчиков SP-01 (США) и компьютерной программы Bioshell. Исследуемые вещества вводили в дозах 30, 50, 100 мкг/кг внутрибрюшинно.
3.1 Изучение острой токсичности исследуемых соединений
Определение острой токсичности селенита натрия, селенита цинка и расчёты LD50 проводили по методу Кербера. Белым мышам (90 особей) массой 20-22 г, обоего пола селенит натрия и селенит цинка вводили в 0,5 мл изотонического раствора внутрибрюшинно в дозах 1, 2, 3, 4, 5, 6, 7, 8 мг/кг. Наблюдение за поведением и состоянием подопытных животных проводилось в течение 7 суток, при этом отмечали изменение внешнего вида, поведенческих реакций и количество погибших животных. Контрольной группе животных (10 особей) вводили физиологический раствор в эквивалентном объеме.
LD50 рассчитывали по формуле:
LD50 = LD100 Σ (Z*D)/m, где
Σ – сумма.
Z – половина суммы числа животных павших от двух последующих доз;
D – интервал между каждыми двумя последующими дозами;
m – число животных на каждую дозу;
За время наблюдения в контрольной группе не погибло ни одного животного, а также не наблюдалось особых изменений во внешнем виде и поведении мышей.
В опытных группах количество погибших животных в зависимости от дозы приведено в таблицах 1 и 2.
Таблица 1 – Влияние различных доз селенита натрия на выживаемость животных
| Доза селенита натрия мг/кг | Общее число животных в группе | Число павших животных | Z | D | Z*D |
| 1 | 6 | 0 | - | - | - |
| 2 | 6 | 0 | 0 | 1 | 0 |
| 3 | 6 | 2 | 1 | 1 | 1 |
| 4 | 6 | 3 | 3.5 | 1 | 3.5 |
| 5 | 6 | 3 | 4.5 | 1 | 4.5 |
| 6 | 6 | 5 | 5.5 | 1 | 5.5 |
| 7 | 6 | 5 | 7.5 | 1 | 7.5 |
| 8 | 6 | 6 | 8 | 1 | 8 |
Отсюда LD50 составила:
LD50 = 8 (1,0+3,5+4,5+5,5+7,5+8,0) / 6 = 3 мг/кг
Результаты проведённых исследований показали, что согласно требованиям табуляции классов токсичности селенит натрия относится ко второму классу токсичности. LD50 составила 3 мг/кг.
Таблица 2 - Влияние различных доз селенита цинка на выживаемость животных
| Доза селенита натрия мг/кг | Общее число животных в группе | Число павших животных | Z | D | Z*D |
| 1 | 6 | 0 | - | - | - |
| 2 | 6 | 0 | 0 | 1 | 0 |
| 3 | 6 | 0 | 0 | 1 | 0 |
| 4 | 6 | 2 | 1 | 1 | 1 |
| 5 | 6 | 4 | 4 | 1 | 4 |
| 6 | 6 | 4 | 6 | 1 | 6 |
| 7 | 6 | 5 | 6.5 | 1 | 6.5 |
| 8 | 6 | 6 | 8 | 1 | 8 |
Отсюда LD50 составила:
LD50 = 8 (1,0+4,0+6,0+6,5+8,0) / 6 = 3,75 мг/кг
Результаты проведённых исследований показали, что согласно требованиям табуляции классов токсичности селенит цинка относится ко второму классу токсичности. LD50 составила 3,75мг/кг.
3.2 Влияние селенита натрия и селенита цинка на мозговой кровоток
Эксперименты проведены на 40 белых беспородных крысах, выращенных в стандартных условиях вивария, массой 200,0-230,0 г. В качестве наркоза использовали хлоралгидрат в дозе 300 мг/кг. Вещества вводили внутрибрюшинно, однократно, в виде изотонического раствора в дозах 100, 50 и 30 мкг/кг. МК регистрировали методом водородного клиренса. Полученные результаты сравнивали с исходными значениями МК.
Внутрибрюшинное введение селенита натрия в дозе 30 мкг/кг (6 опытов) вызывало достоверное снижение МК с первых минут эксперимента. Так, на 5 минуте наблюдения степень снижение МК составила 25%, а на 15 минуте - 41,2% по отношению к исходному уровню. В дальнейшем МК продолжал снижаться и на 60 минуте был ниже исходного уровня на 53,7±15,8% (таблица 3).
Таблица 3 - Динамика изменения скорости мозгового кровотока (МК) у белых крыс при введении селенита натрия в дозе 30 мкг/кг, (M±m, n=6, D%)
| Исходные данные |
МК мл/100г/мин153,0±66,3 |
| Время после введения физиологического раствора | % от исходных данных |
| Через 5 мин | -25,0±19,4* |
| 15 мин | -41,2±17,1* |
| 30 мин | -46,0±10,6* |
| 45 мин | -52,1±13,7* |
| 60 мин | -53,7±15,8* |
Примечание - достоверно относительно исходных данных, * р≤0,05
Аналогично изменялся МК после введения селенита натрия в дозе 50 мкг/кг (6 опытов), однако степень его снижения была менее выраженной. На 5 минуте наблюдения МК был ниже исходного уровня на 28,1±13,1%, а к концу наблюдения – на 41,3±10,5% (таблица 4).
Таблица 4 - Динамика изменения скорости мозгового кровотока (МК) у белых крыс при введении селенита натрия в дозе 50 мкг/кг, (M±m, n=6, D%)
| Исходные данные |
МК мл/100г/мин 114,4±30,4 |
| Время после введения физиологического раствора | % от исходных данных |
| Через 5 мин | -28,1±13,1* |
| 15 мин | -28,4±13,7 |
| 30 мин | -25,3±24,6 |
| 45 мин | -32,0±8,1* |
| 60 мин | -41,3±10,5* |
Примечание - достоверно относительно исходных данных, * р≤0,05
При введении селенита натрия в дозе 100 мкг/кг (8 опытов) эффект также наступал с первых минут наблюдения, при этом снижение МК было достоверно ниже чем при введении селенита натрия в дозах 30 и 50 мкг/кг. Так, в ранние сроки наблюдения мозговой кровоток уменьшался незначительно (на 6-14%). К концу экспериментов на 60 минуте МК был меньше исходного уровня на 20,5±12,7% (таблица 5).
Таблица 5 - Динамика изменения скорости мозгового кровотока (МК) у белых крыс при введении селенита натрия в дозе 100 мкг/кг, (M±m, n=8, D%)
| Исходные данные |
МК мл/100г/мин184,6±59,6 |
| Время после введения физиологического раствора | % от исходных данных |
| Через 5 мин | -6,6±4,6* |
| 15 мин | -13,4±6,6* |
| 30 мин | -16,2±15,9 |
| 45 мин | -22,0±11,1* |
| 60 мин | -20,5±12,7* |
Примечание - достоверно относительно исходных данных, * р≤0,05
Суммарные результаты опытов представлены на рисунке 1.
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
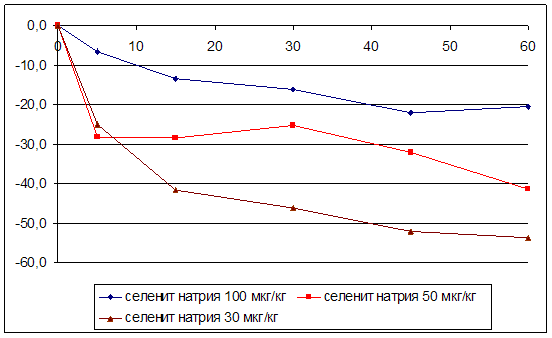
Рисунок 1 - Динамика изменения МК у белых крыс при введении селенита натрия в дозах 30 мкг/кг, 50 мкг/кг, 100 мкг/кг
Примечание - достоверно относительно исходных данных, * р≤0,05
По оси абсцисс время наблюдения в минутах; по оси ординат – изменение мозгового кровотока в процентах от исходного уровня
Внутрибрюшинное введение селенита цинка в дозе 30 мкг/кг (6 опытов) вызывало достоверное снижение МК с первых минут эксперимента. Так, на 5 минуте наблюдения степень снижения МК составила 18,3%, по отношению к исходному уровню. В дальнейшем МК также сохранял тенденцию к снижению. К 30 минуте наблюдения МК был ниже исходного уровня в среднем на 39,8%. К 45 минуте МК опытных животных стабилизировался. Снижение по отношению к исходному уровню составило 38,6%. На 60 минуте наблюдения МК остался ниже исходного уровня на 39,9% (таблица 6).
Таблица 6 - Динамика изменения скорости мозгового кровотока (МК) у белых крыс при введении селенита цинка в дозе 30 мкг/кг, (M±m, n=6, D%)
| Исходные данные |
МК мл/100г/мин149,9±27,1 |
| Время после введения физиологического раствора | % от исходных данных |
| Через 5 мин | -18,3±17,3* |
| 15 мин | -24,3±10,9* |
| 30 мин | -39,8±9,8* |
| 45 мин | -38,6±15,3* |
| 60 мин | -39,9±22,2* |
Примечание - достоверно относительно исходных данных, * р≤0,05
В опытах с внутрибрюшинным введением селенита цинка в дозе 50 мкг/кг также отмечалась тенденция к снижению МК. Однако достоверно МК снижался только на 45 минуте, степень его снижения составила в среднем 38,2% (таблица 7).
Таблица 7 - Динамика изменения скорости мозгового кровотока (МК) у белых крыс при введении селенита цинка в дозе 50 мкг/кг, (M±m, n=6, D%)
| Исходные данные |
МК мл/100г/мин86,7±19,1 |
| Время после введения физиологического раствора | % от исходных данных |
| Через 5 мин | -14,6±22,4 |
| 15 мин | -29,5±21,2 |
| 30 мин | -32,1±27,2 |
| 45 мин | -38,2±21,5* |
| 60 мин | -39,8±27,5 |
Примечание - достоверно относительно исходных данных, * р≤0,05
При введении селенита цинка в дозе 100 мкг/кг (8 опытов) выраженный вазоконстрикторный эффект наступал с первых минут наблюдения, при этом степень снижения МК к концу наблюдения была ниже, чем при введении селенита цинка в дозах 30 и 50 мкг/кг. Так, на 5 минуте наблюдения отмечалось достоверное снижение МК на 24,0±18,3%. Наиболее выраженная вазоконстрикторная реакция наблюдалась на 15 минуте, когда МК был в среднем на 30,7% ниже исходного уровня. В дальнейшем величина МК не изменялась (таблица 8).
Таблица 8 - Динамика изменения скорости мозгового кровотока (МК) у белых крыс при введении селенита цинка в дозе 100 мкг/кг, (M±m, n=8, D%)
| Исходные данные |
МК мл/100г/мин101,2±22,3 |
| Время после введения физиологического раствора | % от исходных данных |
| Через 5 мин | -24,0±18,3* |
| 15 мин | -30,7±17,1* |
| 30 мин | -29,1±13,2* |
| 45 мин | -25,4±17,3* |
| 60 мин | -27,4±15,0* |
Примечание - достоверно относительно исходных данных, * р≤0,05
Данные о влиянии различных доз селенита цинка на мозговой кровоток представлены на рисунке 2.
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
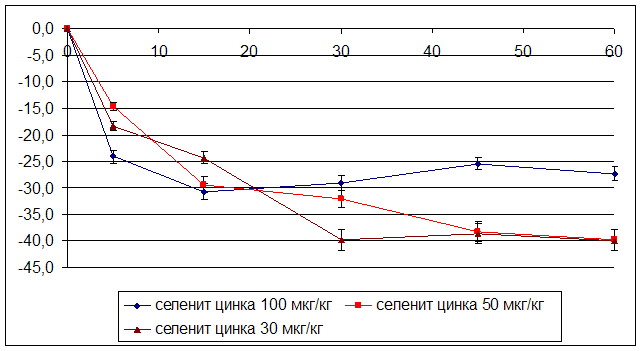
Примечание - достоверно относительно исходных данных, * - р≤0,05
По оси абсцисс время наблюдения в минутах; по оси ординат – изменение мозгового кровотока в процентах от исходного уровня
Рисунок 2 - Динамика изменения МК у белых крыс при введении селенита цинка в дозах 30мкг/кг, 50 мкг/кг, 100 мкг/кг
3.3 Влияние селенита натрия и селенита цинка на артериальное давление и частоту сердечных сокращений у бодрствующих крыс
Известно, что снижение уровня артериального давления, а следовательно, и перфузионнного давления у больных с ишемическим инсультом, вызывает дальнейшее ухудшение ишемизированных областей мозга. Поэтому нейропротекторы не должны обладать выраженным гипотензивным эффектом и не вызывать существенного изменения ЧСС. В связи с этим следующим этапом нашей работы было изучение влияния исследуемых веществ на САД и ЧСС.
Эксперименты проведены на 12 бодрствующих нормотензивных крысах обоего пола, массой 220 - 250г. Предварительно за 24 - 48 часов до начала эксперимента под хлоралгидратным наркозом (300 мг/кг, внутрибрюшинно) имплантировали полиэтиленовый катетер в сонную артерию. Регистрацию данных у бодрствующих крыс производили с использованием одноразовых датчиков SP-01 (США) и компьютерной программы “Bioshell 1.00” в реальном масштабе времени на базе персонального компьютера IBM AT 486 [202]. Длительность регистрации показателей составляла 60 минут с момента введения исследуемого вещества. Исследуемые соединения вводили внутрибрюшинно, однократно в дозе 100 мкг/кг, поскольку в этой дозе в меньшей степени снижался мозговой кровоток.
Внутрибрюшинное введение селенита натрия и селенита цинка в дозе 100 мкг/кг (6 опытов) не вызывало достоверного изменения АД по сравнению с исходным уровнем у бодрствующих животных в течение всего срока наблюдения. Однако необходимо отметить, что при введении селенита натрия отмечалась некоторая тенденция к снижению АД на 6%, а при введении селенита цинка - незначительное увеличение АД в среднем на 6%.
Результаты экспериментов представлены на рисунке 3 и в таблицах 9, 10.
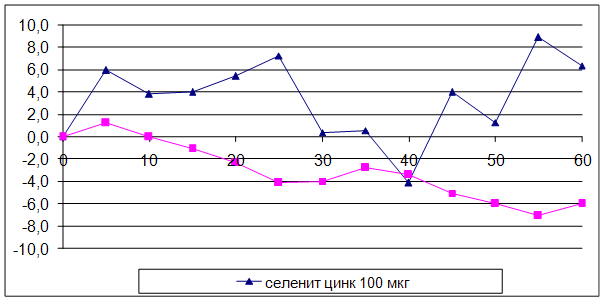
Примечание - достоверно относительно исходных данных, * - р≤0,05
По оси абсцисс время наблюдения в минутах; по оси ординат – артериального давления в процентах от исходного уровня
Рисунок 3 – Влияние селенита натрия и селенита цинка (100 мкг/кг) на системное АД у бодрствующих крыс (M±m, n=6, D%)
Таблица 9 - Динамика изменения АД у белых крыс при введении селенита натрия в дозе 100 мкг/кг, (M±m, n=6, D%) в бодрствующем состоянии
| Исходные данные мм. рт. ст. | 108,7±13,6 | |
| Время после введения вещества | % от исходных данных | Абсолютные значения |
| Через 5 мин | 1,2±6,7 | 110,0±18,6 |
| 10 мин | 0,0±5,7 | 108,7±24,1 |
| 15 мин | -1,1±5,9 | 107,5±27,6 |
| 20 мин | -2,3±7,1 | 106,2±21,1 |
| 25 мин | -4,2±6,9 | 104,2±20,7 |
| 30 мин | -4,0±7,3 | 104,3±24,2 |
| 35 мин | -2,8±7,6 | 105,7±30,1 |
| 40 мин | -3,4±9,5 | 105,0±26,8 |
| 45 мин | -5,1±10,8 | 103,2±20,1 |
| 50 мин | -6,0±11,6 | 102,2±22,9 |
| 55 мин | -7,1±11,6 | 101,0±18,1 |
| 60 мин | -6,0±13,8 | 102,2±18,5 |
Примечание - достоверно относительно исходных данных, *- р≤0,05
Таблица 10 - Динамика изменения АД у белых крыс при введении селенита цинка в дозе 100 мкг/кг, (M±m, n=6, D%) в бодрствующем состоянии
| Исходные данные мм. рт. ст. | 92,0±20,8 | |
| Время после введения вещества | % от исходных данных | Абсолютные значения |
| Через 5 мин | 6,0±19,1 | 97,5±18,6 |
| 10 мин | 3,8±25,2 | 95,5±24,1 |
| 15 мин | 4,0±28,9 | 95,7±27,6 |
| 20 мин | 5,4±21,7 | 97,0±21,1 |
| 25 мин | 7,2±20,9 | 98,7±20,7 |
| 30 мин | 0,4±26,2 | 92,3±24,2 |
| 35 мин | 0,5±32,6 | 92,5±30,1 |
| 40 мин | -4,2±30,4 | 88,2±26,8 |
| 45 мин | 4,0±21,0 | 95,7±20,1 |
| 50 мин | 1,3±24,6 | 93,2±22,9 |
| 55 мин | 8,9±18,0 | 100,2±18,1 |
| 60 мин | 6,3±19,0 | 97,8±18,5 |
Примечание - достоверно относительно исходных данных, * - р≤0,05
Внутрибрюшинное введение селенита натрия и селенита цинка в дозе 100 мкг/кг (6 опытов) не вызывало достоверного изменения ЧСС по сравнению с исходным уровнем у бодрствующих животных в течение всего срока наблюдения. Однако необходимо отметить, что при введении селенитов отмечалась тенденция к некоторому увеличению ЧСС в среднем на 6%.
Результаты экспериментов представлены в таблицах 11, 12.
Таблица 11 - Динамика изменения ЧСС у белых бодрствующих крыс при введении селенита натрия в дозе 100 мкг/кг, (M±m, n=6, D%)
| Исходные данные уд/мин | 392,3±39,5 | |
| Время после введения вещества | % от исходных данных | Абсолютные значения |
| Через 5 мин | 2,1±11,2 | 400,5±45,0 |
| 10 мин | 2,6±11,0 | 402,3±44,1 |
| 15 мин | 1,3±10,6 | 397,5±42,1 |
| 20 мин | 5,7±10,3 | 414,7±42,8 |
| 25 мин | 4,6±9,8 | 410,5±40,4 |
| 30 мин | 6,1±11,1 | 416,2±46,0 |
| 35 мин | 5,1±10,9 | 412,3±44,9 |
| 40 мин | 3,3±10,4 | 405,2±42,2 |
| 45 мин | 4,3±9,9 | 409,0±40,4 |
| 50 мин | 4,0±10,2 | 408,2±41,7 |
| 55 мин | 5,0±9,2 | 411,8±37,8 |
| 60 мин | 2,8±10,6 | 403,3±42,7 |
Примечание - достоверно относительно исходных данных, * - р≤0,05
Таблица 12 - Динамика изменения ЧСС у белых бодрствующих крыс при введении селенита цинка в дозе 100 мкг/кг, (M±m, n=6, D%)
| Исходные данные уд/мин | 363,0±52,3 | |
| Время после введения вещества | % от исходных данных | Абсолютные значения |
| Через 5 мин | 4,5±11,9 | 379,5±45,1 |
| 10 мин | 6,9±12,6 | 388,0±49,1 |
| 15 мин | 8,2±10,1 | 392,8±39,7 |
| 20 мин | 5,7±6,6 | 383,8±25,3 |
| 25 мин | 11,0±12,6 | 402,8±50,8 |
| 30 мин | 5,6±15,9 | 383,3±60,9 |
| 35 мин | 0,4±18,7 | 364,5±68,2 |
| 40 мин | 2,2±15,2 | 370,8±56,2 |
| 45 мин | 5,9±13,4 | 384,3±51,3 |
| 50 мин | 4,0±11,8 | 377,7±44,4 |
| 55 мин | 3,9±5,7 | 377,2±21,5 |
| 60 мин | 6,1±7,4 | 385,0±28,7 |
Примечание - достоверно относительно исходных данных, * - р≤0,05
Выводы по главе 3:
1. Результаты изучения острой токсичности показали, что селенит натрия при введении в брюшную полость по К.К. Сидорову относится ко второму классу токсичности.
2. Селенит цинка по классификации токсичности веществ при введении в брюшную полость по К.К. Сидорову относится ко второму классу токсичности.
3. Селенит натрия и селенит цинка в дозах 30 мкг/кг, 50 мкг/кг и 100 мкг/кг достоверно снижают уровень МК у наркотизированных интактных крыс. С увеличением дозы исследуемых соединений степень снижения МК уменьшается.
4. Селенит натрия и селенит цинка не оказывают существенного влияния на АД и ЧСС бодрствующих белых крыс в дозе 100 мкг/кг.
ГЛАВА 4 ВЛИЯНИЕ СЕЛЕНИТА НАТРИЯ И СЕЛЕНИТА ЦИНКА НА ПСИХОНЕВРОЛОГИЧЕСКИЙ СТАТУС ЖИВОТНЫХ В УСЛОВИЯХ ЭКСПЕРИМЕНТАЛЬНОЙ НОРМЫ
Одним из проявлений сосудистой патологии мозга является нарушение психоневрологического статуса. Для его восстановления используют ноотропные препараты. Однако вещества применяемые в настоящее время для лечения неврологического дефицита, имеют ряд ограничений и недостатков. Поэтому весьма актуальным представляется изучить вопрос о возможности коррекции нарушений поведения, памяти, эмоционального статуса, тревожных расстройств с помощью селенита натрия и селенита цинка. На первом этапе исследования изучали ориентировочно-исследовательскую, двигательную и эмоциональную активность, мнестические функции и координацию движений у интактных животных.
Опыты выполнены на 18 белых крысах обоего пола массой 200 – 250 г. Селенит натрия и селенит цинка вводили в виде изотонического раствора внутрибрюшинно за 50 - 60 минут до проведения эксперимента в дозе 100 мкг/кг. В контрольных опытах, в тех же условиях, вводили физиологический раствор. В каждой группе животных проводилось тестирование психоневрологического статуса. В течение 3-х минут пребывания крысы в «открытом поле» регистрировали двигательную активность (число пересечённых секторов), ориентировочно-исследовательскую активность (вставание на задние лапы) и эмоциональный статус (количество стереотипных движений – груминг, фекальных болюсов).
Исследование проводили на 2, 4 и затем на 7 сутки после первой инъекции исследуемых веществ. Это позволило следить за динамикой изменений психоневрологического статуса ишемизированных животных. Эсперименталь-ные данные статически обработаны и представлены в таблицах 13 – 15.
Таблица 13 – Показатели психоневрологического статуса животных в контрольной серии опытов
|
Время эксперимента (сутки) |
Исследуемые вещества |
Кол-во пройденных секторов |
Кол-во стоек |
Дефекация |
Число уринаций |
Груминг |
Кол-во пройденных центральных секторов |
| 2 день | контроль | 16,0 ± 1,6 | 6,8 ± 1,2 | 3,7 ± 1,4 | 0,3 ± 0,3 | 0,5 ± 0,5 | 0,7 ± 0,3 |
| 4 день | контроль | 13,5 ± 2,4 | 4,3 ± 1,0 | 2,0 ± 1,4 | 0,5 ± 0,3 | 1,8 ± 0,5 | 0,3 ± 0,3 |
| 7 день | контроль | 15,7 ± 2,0 | 6,8 ± 1,1 | 1,5 ± 1,0 | 0,0 ± 0,0 | 1,7 ± 0,6 | 0,7 ± 0,2 |
Таблица 14 – Показатели психоневрологического статуса в группе животных с предварительным введением селенита натрия в дозе 100 мкг/кг
|
Время эксперимента (сутки) |
Исследуемые вещества |
Кол-во пройденных секторов |
Кол-во стоек |
Дефекация |
Число уринаций |
Груминг |
Кол-во пройденных центральных секторов |
| 2 день | Селенит натрия | 15,7 ± 1,2 | 5,2 ± 0,6 | 2,7 ± 0,6 | 0,5 ± 0,3 | 1,0 ± 0,4 | 0,3 ± 0,2 |
| 4 день | Селенит натрия | 14,8 ± 1,2 | 4,7 ± 1,1 | 2,5 ± 0,8 | 0,3 ± 0,2 | 0,8 ± 0,4 | 0,5 ± 0,3 |
| 7 день | Селенит натрия | 16,2 ± 1,1 | 5,7 ± 0,9 | 1,8 ± 0,7 | 0,3 ± 0,2 | 1,3 ± 0,6 | 0,3 ± 0,2 |
Примечание - достоверно относительно контрольных данных, * р≤0,05
Таблица 15 – Показатели психоневрологического статуса животных с предварительным введением селенита цинка в дозе 100 мкг/кг
|
Время эксперимента (сутки) |
Исследуемые вещества |
Кол-во пройденных секторов |
Кол-во стоек |
Дефекация |
Число уринаций |
Груминг |
Кол-во пройденных центральных секторов |
| 2 день | Селенит цинка | 15,8 ± 1,6 | 7,0 ± 0,9 | 3,3 ± 0,6 | 0,8 ± 0,3 | 1,5 ± 0,6 | 0,7 ± 0,4 |
| 4 день | Селенит цинка | 15,0 ± 1,9 | 4,5 ± 1,0 | 2,3 ± 0,8 | 0,5 ± 0,3 | 1,2 ± 0,5 | 0,7 ± 0,3 |
| 7 день | Селенит цинка | 16,0 ± 2,1 | 6,3 ± 0,8 | 2,0 ± 0,6 | 0,2 ± 0,2 | 0,8 ± 0,3 | 0,7 ± 0,3 |
Примечание - достоверно относительно контрольных данных, * р≤0,05
Результаты проведённых исследований не выявили существенных отличий между данными контрольной серии опытов и в тестах с предварительным введением селенита натрия и селенита цинка. В динамике изменений во всех экспериментальных группах отмечается тенденция к снижению уровня эмоциональной тревожности, при этом уровень двигательной и познавательной активности варьировал незначительно, однако также несколько снижался. Данный эффект по-видимому связан с привыканием животных к обстановке эксперимента.
4.2 Изучение влияния исследуемых соединений на процессы памяти и обучения
Опыты проведены на 18 белых крысах обоего пола массой 200 – 250 г. Селенит натрия и селенит цинка вводили внутрибрюшинно за 50 - 60 минут до проведения эксперимента в дозе 100 мкг/кг. В контрольных опытах в тех же условиях вводили физиологический раствор. Обучение животных проводили за 24 часа до первой инъекции исследуемых веществ. В каждой группе из 6 животных проводили тест на воспроизведение УРПИ. При помещении животных в экспериментальную установку регистрировали латентный период захода (ЛПЗ) крысы в тёмную камеру и время пребывания в этой камере в течение 3 минут. Для изучения влияния селенита натрия и селенита цинка на мнестические функции в динамике исследование проводили на 2, 4 и затем на 7 сутки после первой инъекции исследуемых веществ. Полученные экспериментальные данные статически обработаны и представлены в таблицах 16 и 17.
Таблица 16 – Оценка ЛПЗ у белых крыс
| Исследуемые вещества |
Дозы, мкг/кг |
Кол-во животных |
Латентный период первого захода крыс в тёмный отсек в сек. | ||
| 2 день | 4 день | 7 день | |||
| Физ. раствор | 6 | 128,3±21,7 | 115,0±18,3 | 120,8±15,8 | |
| Селенит натрия | 100 | 6 | 122,5±20,0 | 117,5±28,3 | 124,2±37,2 |
| Селенит цинка | 100 | 6 | 119,2±22,8 | 120,2±17,5 | 117,5±25,8 |
Примечание - достоверно относительно контрольных данных, * р≤0,05
Таблица 17 – Оценка времени пребывания белых крыс в тёмном отсеке
| Исследуемые вещества |
Дозы, мкг/кг |
Кол-во животных |
Время пребывания крыс в тёмном отсеке в сек. | |||
| 2 день | 4 день | 7 день |
|
|||
| Физ. раствор | 6 | 19,2±7,5 | 20,8±7,8 | 16,7±8,3 | ||
| Селенит натрия | 100 | 6 | 18,3±8,3 | 20,0±11,7 | 15,0±11,7 | |
| Селенит цинка | 100 | 6 | 20,0±11,7 | 20,8±11,1 | 17,5±10,0 | |
Примечание - достоверно относительно контрольных данных, * р≤0,05
Полученные результаты свидетельствуют о том, что исследуемые соединения в условиях экспериментальной нормы не оказывают существенного влияния на процессы памяти и обучения белых крыс.
4.3 Влияние селенита натрия и селенита цинка на координацию движений
В эксперименте были использованы белые крысы обоего пола массой 200 – 250 г. Селенит натрия и селенит цинка вводили внутрибрюшинно в дозе 100 мкг/кг за 60 минут до проведения эксперимента. В контрольных опытах в тех же условиях вводили физиологический раствор. В каждой группе животных проводилась оценка координации движений.
Крыс помещали на горизонтальный стержень диаметром 4 см, вращающийся со скоростью 3 об/мин. Неспособность животных удерживать равновесие на стержне в течение трёх минут рассматривали как проявление нарушения координации движений. Для изучения координации движений в динамике исследование проводили на 2, 4 и затем на 7 сутки после первой инъекции исследуемых веществ.
Полученные экспериментальные данные статически обработаны и представлены в таблице 18.
Таблица 18 – Влияние селенита натрия и селенита цинка (100 мкг/кг) на координацию движений белых крыс
|
Исследуемые вещества |
Дозы, мкг/кг |
Кол-во животных |
Кол-во животных оставшихся на стержне | |||||
| 2 день | 4 день | 7 день | ||||||
| Абс. | % | Абс. | % | Абс. | % | |||
| Физ. раствор | 8 | 4 | 50,0 | 5 | 62,5 | 5 | 62,5 | |
| Селенит натрия | 100 | 8 | 5 | 62,5 | 6 | 75,0 | 6 | 75,0 |
| Селенит цинка | 100 | 8 | 4 | 50,0 | 5 | 62,5 | 5 | 62,5 |
Примечание - достоверно относительно контрольных данных, * р≤0,05
Результаты проведённых исследований показали, что селенит натрия и селенит цинка не оказывают существенного влияния на координацию движений интактных животных.
Выводы по главе 4
1. В тесте «открытое поле» селенит натрия и селенит цинка не вызывают существенных изменений двигательной, ориентировочно-исследовательской и эмоциональной активности животных.
2. В условиях теста УРПИ установлено, что изучаемые соединения не влияют на процессы обучения и памяти у интактных животных.
3. Селенит натрия и селенит цинка не влияют на координацию движений белых крыс в условиях экспериментальной нормы.
ГЛАВА 5 ИЗУЧЕНИЕ ПРОТИВОГИПОКСИЧЕСКОЙ АКТИВНОСТИ ИССЛЕДУЕМЫХ СОЕДИНЕНИЙ И ИХ ЭФФЕКТИВНОСТИ В УСЛОВИЯХ ЭКСПЕРИМЕНТАЛЬНЫХ ЦЕРЕБРОВАСКУЛЯРНЫХ РАССТРОЙСТВ
В патогенезе органических повреждений головного мозга ведущее значение принадлежит циркуляторным расстройствам. При ишемии головного мозга вследствие уменьшения кровотока ограничивается поступление в ткань кислорода. Происходит нарушение дыхательной цепи митохондрий с повышением уровня восстановленности её компонентов. При этом резко возрастает поток свободных электронов, приводящий к образованию АКФ. В дальнейшем при углублении ишемии нарушаются все жизненно важные функции клетки, усиливаются процессы свободнорадикального окисления, развивается состояние «оксидантного стресса».
Представляло интерес изучить влияние исследуемых соединений на выживаемость животных в условиях нормобарической гипоксии с гиперкапнией, а также на устойчивость белых крыс к циркуляторной гипоксии, вызванной билатеральной окклюзией общих сонных артерий и критическими гравитационными перегрузками.
Учитывая наличие выраженного антиоксидантного действия у селенитов следующим этапом нашего исследования было изучение влияния исследуемых соединений на динамику развития постишемических цереброваскулярных феноменов.
5.1 Влияние селенита натрия и селенита цинка на устойчивость животных к гипоксической гипоксии
Эксперименты проведены на 70 белых беспородных мышах обоего пола массой 23 - 25 г, выращенных в стандартных условиях вивария. Критерием эффективности антигипоксического действия была продолжительность жизни животных. Исследуемые вещества вводили однократно внутрибрюшинно за 10 – 15 минут до начала эксперимента в дозах 30, 50 и 100 мкг/кг. Контролем служил физиологический раствор, вводимый в эквивалентном объеме. Статистика проводилась с помощью «критерия ранговых знаков Уилкоксона» и подтверждена статистикой «критерием знаков» (таблица 19).
Таблица 19 - Влияние селенита натрия и селенита цинка на продолжительность жизни мышей в условиях нормобарической гипоксии (M±m, D%)
| Исследуемые вещества |
Дозы, мкг/кг |
Кол-во животных |
Среднее время жизни | |
| минуты | D% от контроля | |||
| Физ. раствор | 10 | 57,0±9,0 | --- | |
| Селенит натрия | 30 | 10 | 69,5±22,3* | +21,9%* |
| 50 | 10 | 64,5±8,5** | +13,2%** | |
| 100 | 10 | 58,5±7,5 | +2,6% | |
| Селенит цинка | 30 | 10 | 81,5±24,8* | +43,0%** |
| 50 | 10 | 67,5±17,0* | +18,4%* | |
| 100 | 10 | 59,0±8,8 | +3,5% | |
Примечание
1 * - достоверно относительно физ. раствора; р≤0,05
2 ** - достоверно относительно физ. раствора; р≤0,01
Полученные результаты исследований позволяют утверждать, что селенит натрия и селенит цинка достоверно увеличивают продолжительность жизни животных в условиях нормобарической гипоксии в сравнении с контролем в дозах 30 мкг/кг и 50мкг/кг. Так, при предварительном введении селенита натрия в дозе 30 мкг/кг средняя продолжительность жизни мышей увеличилась на 21,9% по отношению к контролю. В дозе 50 мкг/кг время жизни возросло на 13,2%.При введении селенита натрия в дозе 100 мкг/кг продолжительность жизни белых мышей достоверно не изменялась по сравнению с животным контрольной группы. Более выраженный антигипоксический эффект оказывал селенит цинка. Введение селенита цинка в дозе 30 мкг/кг увеличивало продолжительность жизни животных на 43%. С увеличением дозы антигипоксический эффект селенита цинка уменьшался. Продолжительность жизни белых мышей увеличивалась на 18, 4% при введении селенита цинка в дозе 50 мкг/кг и практически не изменялась при его введении в дозе 100 мкг/кг (рисунок 4).
![]()
![]()
![]()
![]()
![]()
![]()
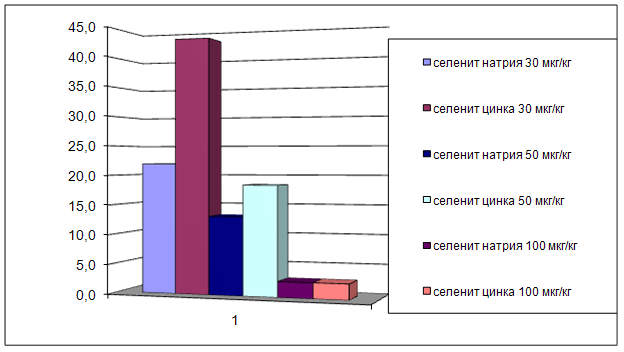
Примечание
1 * - достоверно относительно физ. раствора; р≤0,05
2 ** - достоверно относительно физ. раствора; р≤0,01
Рисунок 4 - Влияние селенита натрия и селенита цинка на продолжительность жизни мышей в условиях нормобарической гипоксии, в процентном отношении к контролю (M±m, D%)
Таким образом, было установлено, что селенит натрия и селенит цинка увеличивают время жизни животных при введении в дозах 50 мкг/кг и 30 мкг/кг.
5.2 Влияние селенита натрия и селенита цинка на устойчивость животных к циркуляторной гипоксии мозга, вызванной билатеральной окклюзией общих сонных артерий
Эксперименты проведены на 24 белых беспородных крысах, обоего пола, массой 220 - 250 г, выращенных в стандартных условиях вивария. Критерием эффективности антигипоксического действия была выживаемость животных в течение 72 часов после двусторонней окклюзии сонных артерий. Исследуемые вещества вводили однократно внутрибрюшинно за 10 – 15 минут до начала эксперимента в дозах 30, 50 и 100 мкг/кг. Контролем служил физиологический раствор в эквивалентном объеме.
Результаты контрольной серии опытов показали, что большинство животных погибло в течение первых суток. Через 72 часа после операции в контрольной группе число выживших животных составило 25%. Нами установлено, что исследуемые соединения обладают антигипоксической активностью.
При профилактическом введении селенита натрия в дозе 30 мкг/кг за 15 минут до ишемии выживаемость животных несколько повысилась. Через 48 часов выживаемость оказалась равной 50%. Введение селенита натрия в дозе 50 мкг/кг вызывало наиболее выраженный антигипоксический эффект. Выживаемость животных на 2-3 сутки составила 62,5%. При увеличении дозы селенита натрия до 100 мкг/кг антигипоксический эффект был равен эффекту при введении соединения в дозе 30 мкг/кг. В опытах с предварительным введением селенита цинка в дозе 30 мкг/кг через 72 часа наблюдения выжило 4 крысы из 8 и выживаемость составила 62,5%. Одинаковый антигипоксический эффект наблюдался при введении селенита цинка в дозах 50 и 100 мкг/кг. Выживаемость животных к концу наблюдения составила 62,5%. Таким образом, полученные результаты свидетельствуют о том, что селенит натрия и селенит цинка обладают антигипоксической активностью, увеличивают процент выживаемости животных при циркуляторной гипоксии, особенно эффективным оказалось введение селенита натрия в дозе 50 мкг/кг. Селенит цинка был активен при его введении в дозах 50 и 100 мкг/кг. Результаты экспериментов представлены в таблице 20 и на рисунке 5.
Таблица 20 - Влияние селенита натрия и селенита цинка на устойчивость белых крыс к циркуляторной гипоксии (M±m, D%)
| Исследуемые вещества |
Дозы, мкг/кг |
Кол-во животных |
Кол-во выживших животных | |||||
|
Через 24 часа |
Через 48 часа |
Через 72 часа |
||||||
| Абс. | % | Абс. | % | Абс. | % | |||
| Физ. раствор | 8 | 5 | 62,5 | 3 | 37,5 | 2 | 25 | |
| Селенит натрия | 30 | 8 | 5 | 62,5 | 4 | 50 | 4 | 50 |
| 50 | 8 | 6 | 75 | 5 | 62,5* | 5 | 62,5* | |
| 100 | 8 | 5 | 62,5 | 4 | 50 | 4 | 50 | |
| Селенит цинка | 30 | 8 | 5 | 62,5 | 5 | 62,5 | 4 | 50 |
| 50 | 8 | 6 | 75 | 6 | 75 | 5 | 62,5* | |
| 100 | 8 | 6 | 75 | 5 | 62,5* | 5 | 62,5* | |
Примечание - достоверно относительно физ. раствора; * - р ≤ 0,05
![]()
![]()
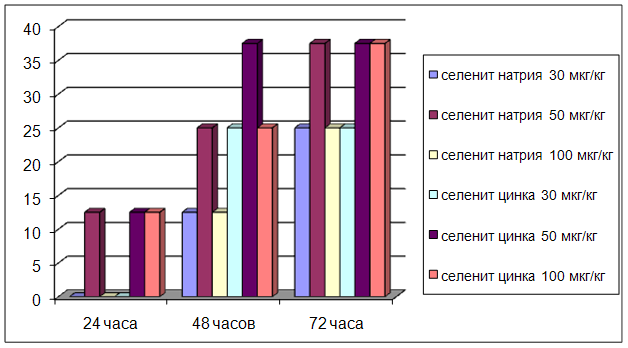
![]()
![]()
![]()
Примечание - достоверно относительно физ. раствора; * - р≤0,05
По оси абсцисс время эксперимента; по оси ординат – выживаемость белых крыс в процентах от исходного уровня
Рисунок 5 - Влияние селенита натрия и селенита цинка на выживаемость белых крыс при циркуляторной гипоксии (M±m, D%)
5.3 Влияние селенита натрия и селенита цинка на устойчивость животных к тотальной ишемии мозга, вызванной критическими гравитационными перегрузками
Учитывая антигипоксическую активность исследуемых соединений при циркуляторной гипоксии мозга, вызванной билатеральной окклюзией общих сонных артерий представляло интерес изучить их действие на устойчивость организма белых крыс к критическим гравитационным перегрузкам.
Известно, что продольные гравитационные перегрузки, создаваемые центрифугой, вызывают нарушения кровоснабжения головного мозга. При краниокаудальном векторе ускорения (положительные радиальные ускорения) происходит перемещение крови в каудальном направлении, в результате чего давление в сосудах мозга снижается и возникает его ишемия, уже при перегрузке 4-5g, давление в сонных артериях снижается до нулевого уровня. Степень проявления цереброваскулярных нарушений зависит от величины и продолжительности перегрузки, а также от градиента её нарастания и спада, что позволяет их дозировать.
Исследования проведены на белых крысах одного возраста и примерно одной массы 220-250г. Белые крысы являются наиболее пригодным объектом для таких исследований, поскольку у них отмечается довольно чёткая зависимость между выживаемостью и величиной перегрузок.
Животных помещали в отдельные продольные ячейки, объём ячеек соответствовал размеру животных и не давал возможности самопроизвольно отклоняться от заданного вектора ускорения. В эксперименте было использовано 36 белых крыс, по 12 в контроле и в опытах с профилактическим введением селенита натрия и селенита цинка. Исследуемые вещества вводились внутрибрюшинно в дозе 100 мкг/кг за 10-15 минут до перегрузок. Исследование устойчивости животных к гравитационным перегрузкам проводили при кранио-каудальном векторе ускорения.
Центрифугирование животных проводили в 2 этапа. На первом этапе исследования крысы подвергались воздействию перегрузок величиной 16 g в течение 10 минут, после чего центрифугу останавливали на 5 минут. На втором этапе исследований, по истечении 5 минутного перерыва, животные подвергались перегрузкам величиной 21 g в течение 5 минут.
В результате исследований было установлено, что большинство животных погибло на втором этапе исследований. Так, в контрольной группе на первой стадии эксперимента погибли 2 крысы из 12, что составило 16,7%, при повторном центрифугировании погибли 3 крысы, что составило 41,7%. При профилактическом введении селенита натрия на первой стадии эксперимента из 12 крыс погибла 1, что составило 8,3 %, на втором этапе погибли 2 крысы, что составило 25%.
Селенит цинка проявлял более выраженный защитный эффект: на первой стадии эксперимента не погибло ни одного животного. Выживаемость крыс составила 100%. На втором этапе исследований погибла 1 крыса из 12, что составило 8,3%. Результаты проведённых исследований статистически обработаны и приведены на рисунке 6 и в таблице 21.
![]()
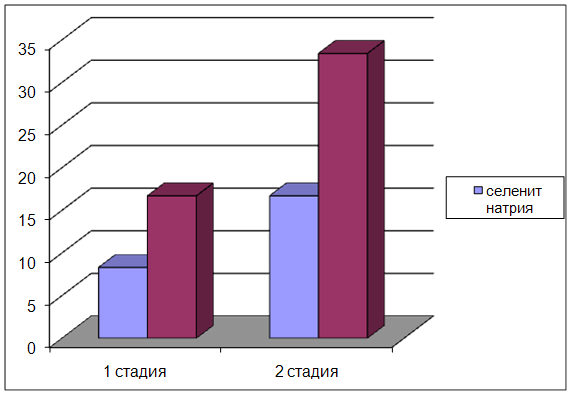
Примечание - достоверно относительно физ. раствора, * - р≤0,05
По оси абсцисс стадия эксперимента; по оси ординат – выживаемость белых крыс в процентах от исходного уровня
Рисунок 6 Влияние селенита натрия и селенита цинка на устойчивость белых крыс к гравитационным перегрузкам в краниокаудальном направлении
Таблица 21 Влияние селенита натрия и селенита цинка (100 мкг/кг, внутрибрюшинно) на устойчивость белых крыс к гравитационным перегрузкам в краниокаудальном направлении
| Вещество | 1 стадия | 2 стадия | ||
|
Количество выживших животных |
% выживших животных |
Количество выживших животных |
% выживших животных |
|
| Контроль | 10 | 83,3% | 7 | 58,3% |
| Селенит натрия | 11 | 91,7% | 9 | 75,0% |
| Селенит цинка | 12 | 100,0% | 11* | 91,7% |
Примечание - достоверно относительно контроля; * - р≤0,05
Таким образом, в результате проведённых исследований было установлено, что селенит натрия и селенит цинка в дозе 100 мкг/кг значительно увеличивают устойчивость животных к гравитационным перегрузкам в краниокаудальном направлении.
5.4 Влияние исследуемых соединений на мозговой кровоток у наркотизированных белых крыс в условиях глобальной ишемии мозга
Эксперименты проведены на белых беспородных крысах обоего пола, наркотизированных хлоралгидратом (300 мг/кг). Ишемию мозга моделировали двухсторонней окклюзией сонных артерий при САД равном 40 мм рт.ст. в течение 10-12 мин.
После ишемии и реинфузии крови в артериальную систему животного осуществляли регистрацию скорости МК методом клиренса водорода на фоне снижения артериального давления. Исследуемые соединения вводились в дозе 100 мкг/кг, внутрибрюшинно.
В контрольных опытах исходное значения МК составляло 101,4 ± 8,4 мл/100г/мин. В течение 15-20 мин после ишемии мозга наблюдалось увеличение МК. На 5 минуте МК достоверно был выше исходного уровня в среднем на 27,8%. К 20 минуте МК восстанавливался и в дальнейшем отмечалось его снижение. На 30 минуте степень снижения МК составила 12,7%, а к концу наблюдения (через 60 мин.) – 37,0%. Таким образом в условиях глобальной ишемии мозга ярко проявляются постишемические цереброваскулярные феномены: фаза реактивной гиперперфузии и фаза гипоперфузии (отсроченного невосстановления кровотока).
Предварительное введение (за 15 мин до ишемии) селенита натрия в дозе 100 мкг/кг вызывало изменение динамики развития цереброваскулярных феноменов в постишемическом периоде (таблица 22 - 24, рисунок 7). Первая фаза становится менее продолжительной и МК увеличивается в меньшей степени. Через 5 минут после ишемии на фоне селенита натрия МК был выше исходного уровня на 8,8%, а в контрольных опытах на 27,8%. Начиная с 15 минуты и по 60 минуту наблюдения отмечалось снижение МК. Однако степень снижения МК на 60 минуте была значительно ниже, чем в контроле.
Результаты исследования с предварительным введением селенита цинка в дозе 100 мкг/кг и последующей ишемией мозга (7 опытов) показали, что в постишемическом периоде фаза гиперперфузии была менее выражена. Так, через 5 минут после ишемии МК был выше исходного уровня на 2,4% (в контроле – 27,8%). Селенит цинка ослаблял фазу гипоперфузии головного мозга. Если в контроле на 60 минуте постишемического периода МК снижался на 37,0%, то под влиянием предварительного введения селенита цинка МК снижался на 19,1%. Таким образом профилактическое введение селенита натрия и селенита цинка существенно изменяет динамику развития постишемических цереброваскулярных феноменов. Приведённые данные о благоприятном влиянии исследуемых соединений на МК в условиях ишемии мозга позволяют в какой-то мере объяснить их защитное действие при циркуляторной гипоксии, вызванной двухсторонней перевязкой общих сонных артерий и критическими гравитационными перегрузками.
Таблица 22 Динамика изменения мозгового кровотока в постишемическом периоде в контрольных опытах
| Исходные данные |
МКмл/100г/мин101,4 ± 8,4 |
| Время после введения физиологического раствора | % от исходных данных |
| Через 5 мин | +27,8 ± 5,1* |
| 15 мин | +3,2 ± 4,6 |
| 30 мин | -12,7 ± 2,1* |
| 45 мин | -20,0 ± 2,5 |
| 60 мин | -37,0 ± 1,9* |
Примечание - достоверно относительно исходных данных, * р≤0,05
Таблица 23 Влияние селенита натрия (100 мкг/кг) на мозговой кровоток белых крыс в постишемическом периоде, (M±m, n=7, D%)
| Исходные данные |
МК мл/100г/мин81,8±27,8 |
| Время после введения селенита натрия | % от исходных данных |
| Через 5 мин | +8,8±29,8* |
| 15 мин | -20,7±30,4* |
| 30 мин | -26,9±41,3* |
| 45 мин | -23,1±30,3* |
| 60 мин | -28,3±41,3* |
Примечание - достоверно относительно исходных данных, * р≤0,05
Таблица 24 Влияние селенита цинка (100 мкг/кг) на мозговой кровоток белых крыс в постишемическом периоде, (M±m, n=7, D%)
| Исходные данные |
МК мл/100г/мин61,6±19,2 |
| Время после введения селенита цинка | % от исходных данных |
| Через 5 мин | +2,4±57,1* |
| 15 мин | +7,2±54,8* |
| 30 мин | -31,8±32,6* |
| 45 мин | -22,6±26,4* |
| 60 мин | -19,1±24,9* |
Примечание - достоверно относительно исходных данных, * р≤0,05
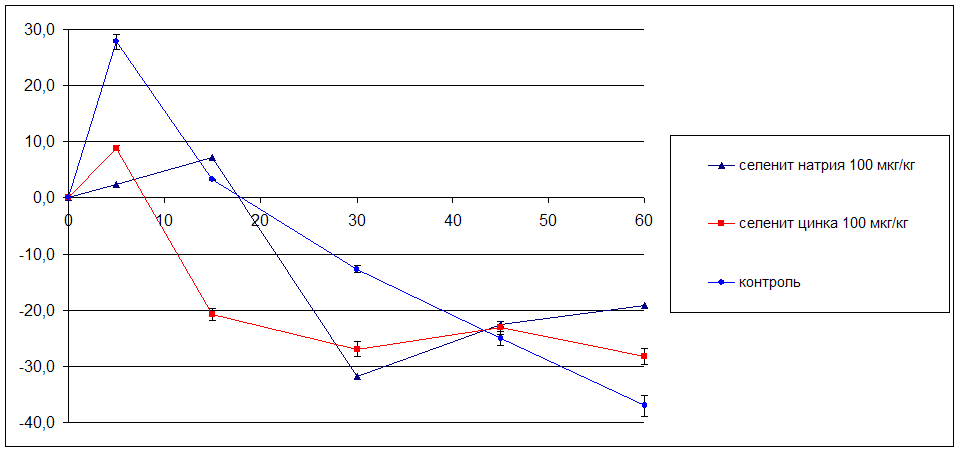
![]()
![]()
![]()
Примечание - достоверно относительно исходных данных, * р≤0,05
По оси абсцисс время наблюдения в минутах; по оси ординат – изменение мозгового кровотока в процентах от исходного уровня
Рисунок 7 Влияние селенита натрия и селенита цинка на динамику развития постишемических цереброваскулярных феноменов
5.5 Влияние селенита натрия и селенита цинка на артериальное давление и сопротивление мозговых сосудов в условиях экспериментальной патологии
Измерение артериального давления у животных проводили с помощью ртутного манометра, соединенного с катетером, предварительно имплантированным в сонную артерию. Для предотвращения свёртывания крови катетер заполняли раствором гепарина. Сопротивление сосудов мозга рассчитывали исходя из данных АД и МК у ишемизированных животных.
Артериальное давление как в контрольной серии экспериментов, так и в группе опытов с предварительным введением исследуемых соединений снижалось с первых минут наблюдения. В контрольной серии снижение АД было значительно меньше и к концу наблюдения составило 5,2%. В группе с предварительным введением селенита натрия (до ишемии мозга) к 60 минуте наблюдения АД снизилось на 19,6%. В опытах с предварительным введением селенита цинка снижение АД было менее выраженным и составило 14,8% к концу наблюдения.
При расчёте ССМ было отмечено, что внутрибрюшинное введение исследуемых соединений оказывало достоверное влияние на сопротивление мозговых сосудов в постишемическом периоде. На протяжении первых 10-15 минут после ишемии отмечалось достоверное снижение сопротивления мозговых сосудов с наиболее выраженным эффектом на 5 минуте на фоне селенита натрия и на 15 минуте - на фоне селенита цинка. К 30 минуте эксперимента ССМ начинало возрастать и на 60 минуте наблюдения было на 12,2% выше исходного уровня при предварительном введении селенита натрия. В меньшей степени к концу наблюдения увеличивалось ССМ. В опытах с предварительным введением селенита цинка степень увеличения ССМ была значительно ниже, чем в контроле (+5,6% и +41,1%, соответственно). Результаты проведённых исследований статистически обработаны и приведены в таблицах 25 - 27 и на рисунке 8.
Таблица 25 – Динамика изменения АД и ССМ у белых крыс в контрольной серии опытов в постишемическом периоде
| Исходные данные |
АДмм рт. ст. |
ССМмм рт. ст./мл/100г/мин |
122,6 ± 3,9 |
1,24 ± 4,0 |
|
| Время после введения физиологического раствора | % от исходных данных | |
| Через 5 мин | -1,1±1,0 | -23,2 ± 4,0* |
| 15 мин | -2,9±1,1* | -3,7 ± 6,0 |
| 30 мин | -7,2 ±2,2* | +5,6 ± 4,6 |
| 45 мин | -6,1±2,4 | +23 ± 5,7 |
| 60 мин | -5,2 ±2,2* | +41,1 ± 9,9* |
Примечание - достоверно относительно исходных данных, * р≤0,05
Таблица 26 - Динамика изменения АД и ССМ у белых крыс в постишемическом периоде получавших селенит натрия в дозе 100 мкг/кг перед ишемией мозга (M±m, n=7, D%)
| Исходные данные |
АД 75,3±8,3 |
|
| Время после введения селенита натрия | % от исходных данных | |
| АД | ССМ | |
| Через 5 мин | -3,8±10,3 | -5,7*±31,9 |
| 15 мин | -9,8*±13,5 | -15,5*±15,8 |
| 30 мин | -9,8*±12,6 | +32,7*±24,5 |
| 45 мин | -17,3*±18,4 | +7,2*±24,3 |
| 60 мин | -14,8*±14,2 | +5,6*±23,3 |
Примечание - достоверно относительно исходных данных, * р≤0,05
Таблица 27 - Динамика изменения АД и ССМ в постишемическом периоде у белых крыс, получавших селенит цинка в дозе 100 мкг/кг перед ишемией мозга (M±m, n=7, D%)
| Исходные данные (мл/100г/мин) |
АД 76,0±10,0 |
|
| Время после введения селенита цинка | % от исходных данных | |
| АД | ССМ | |
| Через 5 мин | -8,4*±9,5 | -15,8*±43,1 |
| 15 мин | -7,8*±10,8 | +16,4*±39,8 |
| 30 мин | -12,0*±13,3 | +20,5*±31,2 |
| 45 мин | -16,9*±11,5 | +8,2*±41,8 |
| 60 мин | -19,6*±9,1 | +12,2*±33,8 |
Примечание - достоверно относительно исходных данных, * р≤0,05
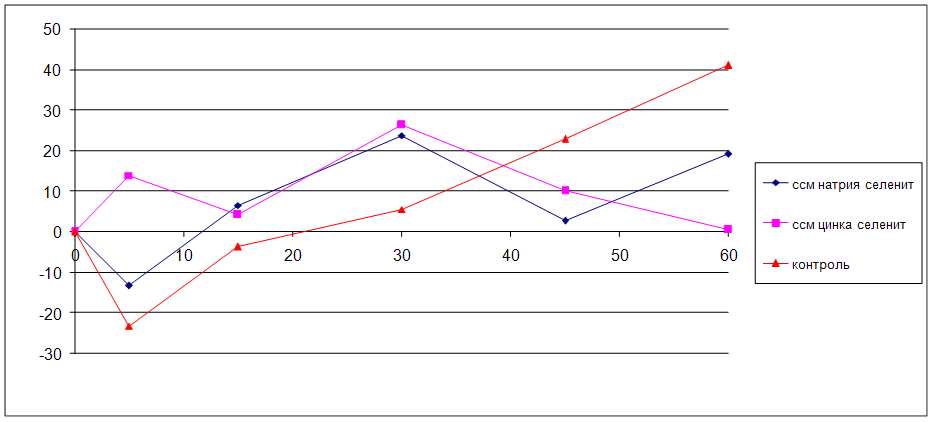
![]()
![]()
Примечание - достоверно относительно исходных данных, * р≤0,05
По оси абсцисс время наблюдения в минутах; по оси ординат – изменение ССМ в процентах от исходного уровня
Рисунок 8 - Динамика изменения ССМ в постишемическом периоде у белых крыс получавших селенит натрия и селенит цинка перед ишемией мозга
Выводы по главе 5
1. Селенит натрия и селенит цинка в дозах 30 и 50 мкг/кг достоверно увеличивают продолжительность жизни животных в условиях нормобарической гипоксии с гиперкапнией.
2. Селенит натрия и селенит цинка увеличивали выживаемость животных в условиях циркуляторной гипоксии мозга в сравнении с контролем.
3. Селенит натрия и селенит цинка в дозе 100 мкг/кг значительно увеличивают устойчивость животных к тотальной ишемии мозга, вызванной критическими гравитационными перегрузками в кранио-каудальном направлении.
4. В условиях экспериментальной ишемии мозга исследуемые соединения при их профилактическом введении существенно изменяют динамику развития постишемических цереброваскулярных феноменов, уменьшают степень проявления реактивной гиперемии и тормозят развитие фазы невосстановления мозгового кровотока.
5. Общая гипотензия, характерная для постишемического периода несколько усиливалась при предварительном введении селенита натрия и селенита цинка. Расчёт ССМ показал, что при введении селенита натрия к 60 минуте наблюдения оно было примерно в 2 раза ниже, чем в контрольной серии опытов, а при введении селенита цинка к концу эксперимента ССМ было близко к исходному.
ГЛАВА 6 ИЗУЧЕНИЕ ПСИХОТРОПНОЙ АКТИВНОСТИ СЕЛЕНИТА НАТРИЯ И СЕЛЕНИТА ЦИНКА В УСЛОВИЯХ ТОТАЛЬНОЙ ИШЕМИИ МОЗГА
При сосудистой патологии мозга в клинике отмечается ряд расстройств его деятельности: нарушение поведения, памяти (в том числе, амнезия) эмоционального статуса, тревожные расстройства. Нарушения в онтогенезе нервно-психических процессов, прежде всего когнитивных, могут проявляться не сразу, а через длительный период после патогенетического воздействия. Однако при оказании помощи пациентам практически все системы лечения направлены на ликвидацию уже развившейся патологии, хотя часто церебральной катастрофе предшествует продолжительный период обратимых нарушений, когда ещё возможно превентивное воздействие. Формирование 50% от окончательного объёма инфаркта происходит в течение первых 90 минут с момента развития инсульта, 80% - в течение 360 минут, в связи с чем первые 3-6 часов получили название «терапевтического окна», внутри которого лечебные мероприятия могут быть наиболее эффективными. Время «доформирования» инфарктных изменений после НМК в каждом случае индивидуально и составляет от 3 до 7 суток [1].
Согласно данным литературы (М.В. Биленко, 1989, В.Б. Семенютин и соавт. 1997) окклюзия артерий мозга сопровождается значительной вариабельностью ишемических повреждений и степенью неврологического дефицита. Известно, что продольные гравитационные перегрузки при краниокаудальном векторе ускорения вызывают нарушения кровоснабжения головного мозга. Степень проявления цереброваскулярных нарушений зависит от величины и продолжительности перегрузки, а также от градиента её нарастания и спада, что позволяет их дозировать. Наряду с изменениями показателей мозговой гемодинамики у животных, подвергшихся воздействию гипергравитации, отмечаются изменения неврологического статуса, поведенческой активности и уровня эмоциональной тревожности.
Известно, что антиоксиданты обладают ноотропными свойствами. Учитывая выраженную антигипоксическую активность селенита натрия и селенита цинка представляло интерес изучить их влияние на обучаемость и процессы памяти, на двигательную, ориентировочно-исследовательскую, эмоциональную активность и координацию движений у животных, подвергшихся тотальной ишемии мозга.
6.1 Изучение влияния селенита натрия и селенита цинка на психоневрологический статус белых крыс, подвергшихся воздействию гравитационных перегрузок (профилактическое введение)
Продольные гравитационные перегрузки создавались центрифугой. В эксперименте использовались белые крысы массой 220-250г, выжившие после перегрузок. Животные были разделены на три исследуемые группы. Контрольную группу составили 7 крыс, получавших изотонический раствор, вторую группу – крысы, которым профилактически до перегрузок вводили селенит натрия в дозе 100 мкг/кг (8 животных). Третьей группе животных (11 крыс) за 10-15 минут до ишемии вводили селенит цинка в дозе 100 мкг/кг. В дальнейшем в каждой группе животных проводилась оценка психоневрологического статуса.
Оценку влияния селенита натрия и селенита цинка на двигательную, ориентировочно-исследовательскую и эмоциональную активность животных проводили в тесте «открытое поле». Установка представляет собой круг диаметром 80 см, с бортиком 20 см, разделённый на 8 радиальных секторов и одним в центре. Тестируемое животное помещали в центральный сектор, спиной к экспериментатору. В течение 3 минут наблюдения регистрировали число пересечённых секторов (двигательная активность), вставание на задние лапы и число пересечённых центральных секторов (ориентировочно-исследовательская активность), количество стереотипных движений (груминг), эмоциональную активность (дефекация, уринация).
Исследование проводили на второй, четвёртый и седьмой день после перенесённых перегрузок. Это позволило следить за динамикой изменений психоневрологического статуса ишемизированных животных.
Результаты проведённых исследований показали, что в постишемическом периоде в контрольных опытах отмечалось понижение двигательной активности, о чём свидетельствует уменьшение числа пересечённых секторов, снижение познавательной активности (уменьшение числа вертикальных стоек) крыс и их эмоционального состояния (уменьшение актов груминга и число дефекаций). Эти нарушения наиболее ярко проявлялись в течение первых дней после ишемии, а в последующие сутки несколько ослабевали. Селенит натрия и селенит цинка при их предварительном до ишемии введении повышали двигательную активность крыс, перенесших ишемию мозга. Более выраженный эффект оказывал селенит цинка, под влиянием которого значительно увеличивалось число пересечённых животными секторов. Профилактическое введение селенита натрия уменьшало у крыс число вертикальных стоек, стереотипных движений и не изменяло число дефекаций по сравнению с контролем, что свидетельствует о некотором снижении исследовательской и эмоциональной активности. Селенит цинка повышал ориентировочно-исследовательскую активность (увеличивал число стоек) и снижал уровень эмоционального состояния (уменьшал акты груминга и дефекаций) крыс в постишемическом периоде.
Полученные экспериментальные данные статически обработаны и представлены в таблицах 28 – 30.
Таблица 28 Показатели психоневрологического статуса контрольной группы животных в постишемическом периоде
|
Время эксперимента (сутки) |
Исследуемые вещества |
Кол-во пройденных секторов |
Кол-во Стоек |
Дефекация |
Число уринаций |
Груминг |
Кол-во пройденных центральных секторов |
| 2 день | физ. раствор | 12,0±2,6 | 7,9±2,7 | 2,4±1,5 | 0,9±0,7 | 2,6±1,1 | 0,0±0,0 |
| 4 день | физ. раствор | 10,6±3,6 | 4,7±2,5 | 2,9±2,1 | 0,7±0,6 | 3,4±0,7 | 0,0±0,0 |
| 7 день | физ. раствор | 13,0±2,6 | 7,0±7,0 | 1,9±1,3 | 0,3±0,5 | 2,3±0,7 | 0,2±0,2 |
Таблица 29 Показатели психоневрологического статуса у животных в постишемическом периоде предварительным введением селенита натрия (100 мкг/кг)
|
Время эксперимента (сутки) |
Исследуемые вещества |
Кол-во пройденных секторов |
Кол-во стоек |
Дефекация |
Число уринаций |
Груминг |
Кол-во пройденных центральных секторов |
| 2 день | Селенит натрия | 10,8±1,9 | 4,1±1,0* | 2,3±1,3 | 0,3±0,6 | 1,0±1,0 | 0,1±0,3 |
| 4 день | Селенит натрия | 12,4±3,3 | 5,4±2,2 | 3,3±2,2 | 1,3±0,3 | 1,3±0,8* | 0,1±0,3 |
| 7 день | Селенит натрия | 14,4±2,0 | 6,4±1,2 | 1,3±1,0 | 0,1±0,3 | 1,6±0,8 | 0,3±0,4 |
Примечание - достоверно относительно контроля, * р≤0,05
Таблица 30 Показатели психоневрологического статуса у животных в постишемическом периоде с предварительным введением селенита цинка (100 мкг/кг)
|
Время эксперимента (сутки) |
Исследуемые вещества |
Кол-во пройденных секторов |
Кол-во стоек |
Дефекация |
Число уринаций |
Груминг |
Кол-во пройденных центральных секторов |
| 2 день | Селенит цинка | 15,4±3,9 | 8,0±3,1 | 1,6±1,1 | 0,3±0,4 | 0,5±0,6* | 0,3±0,4 |
| 4 день | Селенит цинка | 14,2±2,2 | 11,3±2,8* | 2,8±2,3 | 0,5±0,6 | 1,0±0,7* | 0,3±0,4 |
| 7 день | Селенит цинка | 17,3±2,4* | 8,7±2,3 | 1,5±1,2 | 0,2±0,3 | 0,9±0,8* | 0,4±0,5 |
Примечание - достоверно относительно контроля, * р≤0,05
Таким образом, полученные экспериментальные данные свидетельствуют, о том, что однократное введение селенита натрия и селенита цинка до ишемии оказало положительное влияние на организм животных в постишемическом периоде. В опытах с предварительным введением исследуемых веществ уменьшился уровень эмоциональной тревожности, увеличилась двигательная активность, а на фоне селенита цинка повышалась и познавательная активность животных по сравнению с контрольной группой.
Для изучения влияния исследуемых соединений на различные фазы памяти - обучение белых крыс проводили через 24 часа после ишемии в методике условного рефлекса пассивного избегания в модификации Буреш, Бурешова. В каждой группе животных проводили тест на воспроизведение УРПИ. При помещении крыс в экспериментальную установку регистрировали латентный период захода (ЛПЗ) крысы в тёмную камеру и время пребывания (ВП) в этой камере в течение 3 минут. Для изучения ЛПЗ и ВП в тёмном отсеке в динамике - исследование проводили на 2, 4 и затем на 7 сутки после обучения животных.
Поскольку обучение животных проводилось после ишемии, а введение исследуемых соединений проводили до ишемии это позволило судить о степени неврологических нарушений ишемизированных животных. В ходе экспериментов было установлено, что в контрольной группе животных отмечалось ухудшение процессов обучения и процессов воспроизведения памятного следа. Эти эффекты наиболее ярко проявлялись в первые дни наблюдения и несколько ослабевали к седьмому дню наблюдения. Селенит натрия и селенит цинка значительно увеличивали ЛПЗ и ВП. Однако выраженность эффекта селенитов была различной. Наиболее активным оказался селенит цинка. Изучаемые соединения у интактных животных способствовали сохранению навыка УРПИ. Эти данные позволяют сделать вывод, что нарушение памяти крыс, по-видимому, связано с ухудшением процессов фиксации обучения или консолидации памяти (переход кратковременной памяти в долговременную). Полученные экспериментальные данные статистически обработаны и представлены в таблицах 31 - 34.
Таблица 31 Оценка ЛПЗ у белых крыс получавших селенит натрия и до ишемии мозга
| Исследуемые вещества |
Дозы, мкг/кг |
Кол-во животных |
Латентный период первого захода крыс в тёмный отсек в сек. |
|
||
| 2 день | 4 день | 7 день | ||||
| Физ. раствор | 7 | 88,6±30,2 | 122,1±41,0 | 153,6±24,5 |
|
|
| Селенит натрия | 100 | 9 | 120,6±30,7 | 127,8±40,2 | 147,2±34,8 |
|
Примечание - достоверно относительно контроля (интактные крысы), # - р≤0,05
Таблица 32 Оценка ЛПЗ у белых крыс получавших селенит цинка до ишемии мозга
| Исследуемые вещества |
Дозы, мкг/кг |
Кол-во животных |
Латентный период первого захода крыс в тёмный отсек в сек. |
|
||
| 2 день | 4 день | 7 день | ||||
| Физ. раствор | 7 | 88,6±30,2 | 122,1±41,0 | 153,6±24,5 |
|
|
| Селенит цинка | 100 | 11 | 155,5±31,2 | 145,9±22,6 | 150,0±27,3 |
|
Примечание
1 достоверно относительно контроля (крысы перенесшие ишемию мозга), * - р≤0,05
2 достоверно относительно контроля (интактные крысы), # - р≤0,05
3 достоверно относительно селенита цинка (интактные крысы), $ - р≤0,05
Таблица 33 Оценка времени пребывания белых крыс в тёмном отсеке у белых крыс, получавших селенит натрия до ишемии мозга
| Исследуемые вещества |
Дозы, мкг/кг |
Кол-во животных |
Время пребывания крыс в тёмном отсеке в сек. | ||
| 2 день | 4 день | 7 день | |||
| Физ. раствор | 7 | 20,0±10,0 | 11,4±7,8 | 3,6±4,1 | |
| Селенит натрия | 100 | 9 | 9,4±6,2 | 5,0±5,0 | 4,2±4,0 |
Примечание
1 достоверно относительно контроля (крысы перенесшие ишемию мозга), * - р≤0,05
2 достоверно относительно контроля (интактные крысы), # - р≤0,05
3 достоверно относительно селенита цинка (крысы перенесшие ишемию мозга), $ - р≤0,05
Таблица 34 Оценка времени пребывания белых крыс в тёмном отсеке у белых крыс, получавших селенит цинка до ишемии мозга
| Исследуемые вещества |
Дозы, мкг/кг |
Кол-во животных |
Время пребывания крыс в тёмном отсеке в сек. | ||
| 2 день | 4 день | 7 день | |||
| Физ. раствор | 7 | 20,0±10,0 | 11,4±7,8 | 3,6±4,1 | |
| Селенит цинка | 100 | 11 | 1,8±2,6 | 6,8±5,3 | 4,1±4,5 |
Примечание
1 достоверно относительно контроля (крысы перенесшие ишемию мозга), * - р≤0,05
2 достоверно относительно контроля (интактные крысы), # - р≤0,05
3 достоверно относительно селенита натрия (крысы перенесшие ишемию мозга), $ - р≤0,05
Следующим этапом исследования была оценка координации движений в тесте «вращающийся стержень». Селенит натрия и селенит цинка вводили внутрибрюшинно, однократно, за 10-15 минут до проведения эксперимента в дозе 100 мкг/кг. В контрольных опытах в тех же условиях вводили физиологический раствор.
Крыс помещали на горизонтальный стержень диаметром 4 см, вращающийся со скоростью 3 об/мин. Неспособность животных удерживать равновесие на стержне в течение трёх минут рассматривали как проявление нарушения координации движений. Для изучения координации движений в динамике исследование проводили на 2, 4 и затем на 7 сутки после ишемии. Полученные экспериментальные данные статически обработаны и представлены в таблице 35 и 36.
Таблица 35 Влияние профилактического введения селенита натрия на координацию движений белых крыс перенесших тотальную ишемию мозга
| Исследуемые вещества |
Дозы, мкг/кг |
Кол-во крыс |
Кол-во животных оставшихся на стержне | |||||
| 2 день | 4 день | 7 день | ||||||
| Абс. | % | Абс. | % | Абс. | % | |||
| Физ. раствор | 7 | 3 | 42,9 | 4 | 57,2 | 3 | 42,9 | |
| Селенит натрия | 100 | 9 | 4 | 44,4$ | 3 |
33,3 & |
6 | 66,7 |
Примечание
1 достоверно относительно исходных данных с веществом, интактные крысы, & р≤0,05
2 достоверно относительно селенита цинка (крысы перенесшие ишемию мозга), $ - р≤0,05
Таблица 36 Влияние профилактического введения селенита цинка на координацию движений белых крыс перенесших тотальную ишемию мозга
| Исследуемые вещества |
Дозы, мкг/кг |
Кол-во крыс |
Кол-во животных оставшихся на стержне | |||||
| 2 день | 4 день | 7 день | ||||||
| Абс. | % | Абс. | % | Абс. | % | |||
| Физ. раствор | 7 | 3 | 42,9 | 4 | 57,2 | 3 | 42,9 | |
| Селенит цинка | 100 | 11 | 2 |
18,2 #* |
3 |
27,3 #$ |
6 |
54,5 & |
Примечание
1 достоверно относительно контроля (интактные крысы), # - р≤0,05
2 достоверно относительно селенита натрия (крысы перенесшие ишемию мозга), *- р≤0,05
3 достоверно относительно селенита натрия (интактные крысы), $ - р≤0,05
Проведённые эксперименты показали, что в постишемическом периоде в группе контрольных животных отмечалось ухудшение координации движений. Селенит цинка и селенит натрия значительно ослабляли вызванное ишемией нарушение координации движений, что проявлялось в увеличении времени удержания крыс на вращающемся стержне. Так, в контрольных опытах количество животных, удержавшихся на вращающемся стержне в течение 3-х минут, составило 42,9%. При предварительном введении селенита натрия и селенита цинка значительно повышался процент крыс, удержавшихся на стержне, и к концу недели эти показатели составили соответственно: 62,5% и 54,6%
Таким образом, после ишемии отмечались значительные нарушения координации движений животных. Однократное профилактическое введение селенита натрия или селенита цинка оказывало положительное влияние на данный показатель в постишемическом периоде.
6.2 Изучение терапевтического действия исследуемых соединений на психоневрологический статус животных, подвергшихся воздействию гравитационных перегрузок
Продольные гравитационные перегрузки, величиной 9g в течение 10 мин. создавались центрифугой. В эксперименте использовались белые крысы массой 220-250г. Животные были разделены на три исследуемые группы, по 6 животных в каждой. В контрольной группе – крысы, получавшие изотонический раствор. Вторая и третья группа – крысы, которым через 50 – 60 минут после перегрузок внутрибрюшинно вводили селенит натрия и селенит цинка в дозе 100 мкг/ и затем в течение недели за 30-40 минут до эксперимента.
Оценка влияния селенита натрия и селенита цинка на ориентировочно исследовательскую и двигательную активность животных проводили в тесте «открытое поле». В течение 3 минут наблюдения регистрировали число пересечённых секторов (двигательная активность), вставание на задние лапы и число пересечённых центральных секторов (ориентировочно-исследовательская активность), количество стереотипных движений (груминг), эмоциональную активность (дефекация, уринация).
Исследование проводили на второй, четвёртый и седьмой день после перенесённых перегрузок. Это позволило следить за динамикой изменений психоневрологического статуса ишемизированных животных. Полученные экспериментальные данные статически обработаны и представлены в таблицах 37-39.
Результаты проведённых исследований показали, что в постишемическом периоде в контрольных опытах отмечено понижение двигательной активности, о чём свидетельствует уменьшение числа пересечённых секторов, снижение познавательной активности (уменьшение числа вертикальных стоек) крыс и их эмоционального состояния (уменьшение актов груминга и числа дефекаций).
Таблица 37 – Показатели психоневрологического статуса животных контрольной группы животных в постишемическом периоде
|
Время эксперимента (сутки) |
Исследуемые вещества |
Кол-во пройденных секторов |
Кол-во стоек |
Дефекация |
Число уринаций |
Груминг |
Кол-во пройденных центральных секторов |
| 2 день | физ. раствор | 13,0±4,0 | 5,7±4,3 | 3,0±1,7 | 0,3±0,4 | 0,8±0,6 | 0,3±0,6 |
| 4 день | физ. раствор | 12,2±3,2 | 6,5±2,5 | 3,0±1,0 | 0,3±0,4 | 0,8±0,8 | 0,3±0,4 |
| 7 день | физ. раствор | 11,8±5,4 | 6,0±3,7 | 3,2±1,8 | 0,7±0,7& | 1,0±0,7 | 0,3±0,4 |
Таблица 38 – Влияние селенита натрия (100 мкг) на показатели психоневрологического статуса животных, перенесших тотальную ишемию мозга (лечебного введения)
|
Время эксперимента (сутки) |
Исследуемые вещества |
Кол-во пройденных секторов |
Кол-во стоек |
Дефекация |
Число уринаций |
Груминг |
Кол-во пройденных центральных секторов |
| 2 день | Селенит натрия | 12,5±3,3 | 5,5±2,8 | 2,7±1,8 | 0,8±0,8 |
2,0±1,0 # $ |
0,3±0,4 |
| 4 день | Селенит натрия |
11,5±2,3 * |
5,7±2,9 | 2,5±2,2 | 0,5±0,7 |
0,7±0,7 # |
0,5±0,7 |
| 7 день | Селенит натрия |
10,3±3,3 # $ |
6,5±3,7 | 1,8±1,8 | 0,2±0,3 | 1,2±0,6 | 0,2±0,3 |
Примечание
1 достоверно относительно контроля (интактные крысы), # - р≤0,05
2 достоверно относительно контроля (крысы перенесшие ишемию мозга), $ - р≤0,05
3 достоверно относительно селенита цинка (крысы перенесшие ишемию мозга), *- р≤0,05
Таблица 39 – Влияние селенита цинка (100 мкг) на показатели психоневрологического статуса животных, перенесших тотальную ишемию мозга (лечебного введения)
|
Время эксперимента (сутки) |
Исследуемые вещества |
Кол-во пройденных секторов |
Кол-во стоек |
Дефекация |
Число уринаций |
Груминг |
Кол-во пройденных центральных секторов |
| 2 день | Селенит цинка | 18,7±5,7 | 7,2±4,9 | 3,7±1,8 | 0,8±0,8 | 1,5±1,2 | 1,0±1,0 |
| 4 день | Селенит цинка |
17,0±4,0 * |
7,2±2,9 | 2,7±1,7 | 0,5±0,7 | 1,2±0,9 | 0,7±0,7 |
| 7 день | Селенит цинка |
17,2±4,5 * |
3,5±2,2 # |
2,3±2,3 | 0,7±0,7 | 0,5±0,5 | 0,8±0,8 |
Примечание
1 достоверно относительно контроля (интактные крысы), # - р≤0,05
2 достоверно относительно контроля (крысы перенесшие ишемию мозга), $ - р≤0,05
3 достоверно относительно селенита натрия (крысы перенесшие ишемию мозга), *- р≤0,05
Эти нарушения наиболее ярко проявлялись в течение первых дней после ишемии, а в последующие сутки несколько ослабевали. Селенит натрия и селенит цинка, вводимые после ишемии мозга, повышали двигательную активность крыс. Более выраженный эффект оказывал селенит цинка, под влиянием которого достоверно увеличивалось число пересечённых животными секторов. Лечебное введение селенита натрия уменьшало число вертикальных стоек, стереотипных движений и не изменяло число дефекаций по сравнению с контролем, что свидетельствует о некотором снижении исследовательской и эмоциональной активности. Селенит цинка повышал ориентировочно-исследовательскую активность (увеличивал число стоек) и снижал уровень эмоционального напряжения (уменьшал акты груминга и дефекаций) крыс в постишемическом периоде.
Для выяснения влияния исследуемых соединений на мнестические функции проводили тест на воспроизведение УРПИ. Экспериментальные животные были разделены на 3 группы по 6 животных. Обучение белых крыс осуществляли за 24 часа до ишемии по описанной ранее методике условного рефлекса пассивного избегания. При помещении животных в экспериментальную установку регистрировали латентный период захода (ЛПЗ) крысы в тёмную камеру и время пребывания (ВП) в ней. Для изучения координации движений в динамике исследование проводили на 2, 4 и затем на 7 сутки после ишемии.
Под влиянием ишемии, крысы забывали ситуацию и чаще заходили в тёмный отсек установки, при этом увеличивалось время пребывания в нём. В ходе экспериментов было установлено, что изучаемые соединения у животных, перенесших ишемию мозга, способствовали сохранению навыка УРПИ, что проявлялось в увеличении ЛПЗ и ВП в тёмной камере. Однако выраженность эффектов селенитов была различной. Наиболее активным оказался селенит натрия.
Полученные экспериментальные данные статически обработаны и представлены в таблицах 40 и 41.
Таблица 40 – Оценка ЛПЗ у белых крыс получавших селенит натрия и селенит цинка после ишемии мозга
| Исследуемые вещества |
Дозы, мкг/кг |
Кол-во животных |
Латентный период первого захода крыс в тёмный отсек в сек. | ||
| 2 день | 4 день | 7 день | |||
| Физ. раствор | 6 |
76,7±30,0 # |
85,8±36,1 | 105,8±33,9 | |
| Селенит натрия | 100 | 6 |
83,3±43,9 # |
98,3±54,4 | 110,8±46,1 |
| Селенит цинка | 100 | 6 | 81,7±56,7 | 102,5±42,5 | 112,5±45,8 |
Примечание - достоверно относительно контроля (интактные крысы), # - р≤0,05
Таблица 41 – Оценка ВП в тёмном отсеке белых крыс получавших селенит натрия и селенит цинка после ишемии мозга
| Исследуемые вещества |
Дозы, мкг/кг |
Кол-во животных |
Время пребывания крыс в тёмном отсеке в сек. | ||
| 2 день | 4 день | 7 день | |||
| Физ. раствор | 6 | 15,8±7,8 | 14,2±7,2 | 11,7±6,7 | |
| Селенит натрия | 100 | 6 | 14,2±6,1 | 10,0±6,7# | 7,5±5,8# |
| Селенит цинка | 100 | 6 | 15,0±11,7 | 14,2±9,2 | 10,0±8,3 |
Примечание - достоверно относительно контроля (интактные крысы), # - р≤0,05
Под влиянием ишемии, неврологические нарушения в контрольной группе животных было значительно. Однократное введение селенита натрия или селенита цинка оказывали положительное влияние на «качество» жизни животных в постишемическом периоде. Таким образом, селенит натрия и селенит цинка оказывали положительное влияние на процессы воспроизведения памятного следа в постишемическом периоде.
Для оценки координации движений крыс помещали на горизонтальный стержень диаметром 4 см, вращающийся со скоростью 3 об/мин. Неспособность животных удерживать равновесие на стержне в течение трёх минут рассматривали как проявление нарушения координации движений. Для изучения показателей в динамике исследование проводили на 2, 4 и затем на 7 сутки после ишемии.
Результаты проведённых исследований показали, что в постишемическом периоде в группе контрольных животных отмечалось ухудшение координации движений. Селенит цинка достоверно ослаблял вызванное ишемией нарушение координации движений, что проявлялось в увеличении времени удержания крыс на вращающемся стержне. Так, в контрольных опытах количество животных, удержавшихся на вращающемся стержне в течение 3-х минут, составило 66,7%. При лечебном введении селенита цинка повышался процент крыс, удержавшихся на стержне, и к концу недели эти показатели составил 83,3%. Менее активным оказался селенит натрия, координация движений на его фоне практически не отличалась от контроля.
Полученные экспериментальные данные статически обработаны и представлены в таблицах 42 и 43.
Таблица 42 Тест на координацию движений белых крыс получавших селенит натрия после ишемии мозга
| Исследуемые вещества |
Дозы, мкг/кг |
Кол-во животных |
Кол-во животных оставшихся на стержне | |||||
| 2 день | 4 день | 7 день | ||||||
| Абс. | % | Абс. | % | Абс. | % | |||
| Физ. раствор | 6 | 3 | 50,0 | 3 | 50,0 | 4 | 66,7 | |
| Селенит натрия | 100 | 6 | 3 | 50,0 | 4 | 66,7$ | 4 | 66,7 |
Примечание - достоверно относительно селенита натрия (интактные крысы), $ - р≤0,05
Таблица 43 Тест на координацию движений белых крыс получавших селенит цинка после ишемии мозга
| Исследуемые вещества |
Дозы, мкг/кг |
Кол-во животных |
Кол-во животных оставшихся на стержне | |||||
| 2 день | 4 день | 7 день | ||||||
| Абс. | % | Абс. | % | Абс. | % | |||
| Физ. раствор | 6 | 3 | 50,0 | 3 | 50,0 | 4 | 66,7 | |
| Селенит цинка | 100 | 6 | 2 | 33,3 | 3 | 50,0 | 5 | 83,3 |
Таким образом селениты в оказывали положительное влияние на восстановление координации движений в постишемическом периоде.
Выводы по главе 6
1. Селенит натрия в дозе 100 мкг/кг при профилактическом и лечебном введении повышает горизонтальную и умеренно снижает вертикальную активность, уменьшает уровень эмоционального напряжения белых крыс в постишемическом периоде. Селенит цинка повышает двигательную, ориентировочно-исследовательскую и снижает уровень тревожности крыс перенесших тотальную ишемию мозга при его профилактическом введении.
2. Исследуемые соединения в дозе 100 мкг/кг оказывают существенное влияние на процессы выработки УРПИ: увеличивали длительность латентного периода захода в тёмный отсек и время пребывания в нём. Улучшение мнестических функций отмечалось как при профилактическом, так и при лечебном введении.
3. Селенит натрия и селенит цинка в дозе 100 мкг/кг улучшают координацию движений белых крыс, подвергшихся воздействию критических гравитационных перегрузок, при их профилактическом введении.
ЗАКЛЮЧЕНИЕ
Несмотря на достигнутые успехи практической кардиологии в борьбе с возникновением и последствиями сердечно-сосудистой патологии проблемы фармакологической коррекции кардиологических заболеваний остаётся одной из актуальных проблем медицины. В структуре сердечно-сосудистых заболеваний одно из ведущих мест занимают сосудистые поражения мозга.
В последнее время в практике лечения больных с нарушениями мозгового кровообращения получили широкое распространение антиоксиданты. Применение антиоксидантов используется в терапии ишемического инсульта с целью оптимизации окислительно-востановительных процессов. В проведённых экспериментальных и клинических исследованиях высокую антиоксидантную активность показал селенит натрия.
По данным С.М. Николаева селенит натрия предохраняет миокардиальные клетки от разрушения, ограничивает периинфарктную зону, уменьшает размеры рубцов. М.Д. Савиной и А.Н. Кудриным выявлен антиаритмический эффект селенита натрия на экспериментальных моделях аритмий.
Высокая эффективность селенита натрия связана с его способностью повышать активность GPX. Этот фермент способствует накоплению глютатиона, прекращающего или ограничивающего процессы ПОЛ и его распространение.
Накоплен большой опыт применения селенсодержащих соединений в качестве пищевых добавок. Однако их действие на системное артериальное давление, мозговой кровоток до сих пор не изучено. Учитывая тот факт, что селениты являются выраженными антиоксидантами, представляло интерес изучить их действие на показатели системной гемодинамики и оценить целесообразность применения в качестве нейропротекторов.
Исследуемые соединения – селенит натрия и селенит цинка были синтезированы на кафедре фармацевтической химии ПятГФА, под руководством профессора Е.Н. Вергейчика.
Предметом наших исследований явилось экспериментальное изучение влияния селенита натрия и селенита цинка на объёмную скорость МК, артериальное давление и ЧСС, двигательную активность и эмоциональные реакции крыс в тесте «открытое поле», координацию движений животных и на обучаемость в методике пассивного избегания.
Поскольку исследования, проводимые в нормальных условиях регуляции мозгового кровотока, не всегда отвечают интересам клиники, часть опытов выполнена при экспериментальной патологии. Изучено влияние исследуемых соединений на устойчивость животных к различным видам гипоксий, а также на неврологический статус и кровоснабжение головного мозга в условиях его острой ишемии.
Исследования проведены на 236 белых крысах обоего пола, массой 200 – 250 г, а также на 154 мышах массой 20 - 25 г. В качестве наркоза использовали хлоралгидрат в дозе 300 мг/кг.
Результаты изучения острой токсичности показали, что ЛД50 селенита натрия составляет 3 мг/кг, селенита цинка - 3,75 мг/кг.
Исследуемые соединения в виде водных растворов вводили внутрибрюшинно, в дозах 30, 50, 100 мкг/кг (1/100, 1/60, 1/30 от LD50 для селенита натрия и 1/125, 1/75, 1/38 от LD50 для селенита цинка).
Объёмную скорость мозгового кровотока регистрировали методом водородного клиренса с помощью платинного электрода, расположенного на поверхности сагиттального синуса в области стока синусов.
Регистрацию АД и ЧСС у бодрствующих животных проводили с использованием одноразовых датчиков SP-01 (США) и компьютерной программы Bioshell.
Двигательную активность и эмоциональные реакции крыс изучали в тесте «открытое поле». Вырабатывали условный рефлекс пассивного избегания и регистрировали его воспроизведение. Координацию движений оценивали по времени удержания на вращающемся стержне.
Для создания патологического фона, на котором изучалась эффективность исследуемых соединений, использовались различные модели циркуляторных гипоксий. В одной из них циркуляторную гипоксию воспроизводили двухсторонней перевязкой общих сонных артерий. Критерием эффективности антигипоксического действия была выживаемость животных в течение 72 часов.
Вторую модель ишемии мозга создавали критическими гравитационными перегрузками в кранио-каудальном положении животных.
Нормобарическую гипоксию с гиперкапнией вызывали путём помещения мышей в герметически закрытые банки (т.н. «баночная гипоксия»). Во всех случаях регистрировали время выживания животных.
Для оценки действия исследуемых препаратов на динамику развития постишемических цереброваскулярных феноменов воспроизводили ишемию мозга путем двухсторонней окклюзии сонных артерий в течение 10-12 мин. при снижении САД до 40 мм рт. ст. После ишемии и реинфузии крови в артериальную систему животного осуществляли регистрацию скорости МК методом клиренса водорода.
Результаты наших исследований показали, что под влиянием селенита натрия кровоток в сосудах мозга заметно снижается. На данном рисунке представлена динамика изменения МК после введения селенита натрия в дозах 30, 50, 100 мкг/кг. Плавное снижение МК наблюдается с первых минут введения и продолжается до конца наблюдения во всех изучаемых дозах. Наиболее выраженный эффект наблюдается при введении селенита натрия в дозе 30 мкг/кг. Степень снижения кровотока к 60 минуте составила в среднем 53,7±15,8%.
Аналогичный эффект вызывает селенит цинка. Сразу после введения отмечается стойкое снижение мозгового кровотока. С увеличением дозы селенита цинка степень снижения мозгового кровотока уменьшалась. Так, в дозе 30 мкг/кг на 60 минуте мозговой кровоток был ниже исходного уровня в среднем на 39,9%, а в дозе 100 мкг/кг – на 27,4%.
Известно, что снижение уровня артериального давления, а следовательно, и перфузионнного давления у больных с ишемическим инсультом, вызывает дальнейшее ухудшение ишемизированных областей мозга. Поэтому нейропротекторы не должны обладать выраженным гипотензивным эффектом и не вызывать существенного изменения ЧСС. В связи с этим представляло интерес выяснить действие селенита натрия и селенита цинка на АД И ЧСС.
Наши исследования показали, что селенит натрия и селенит цинка не оказывают существенного влияния на АД и ЧСС у бодрствующих животных. Отмечалось тенденция к снижению АД под влиянием селенита натрия и некоторое увеличение АД при введении селенита цинка. Однако изменения были недостоверными.
При оценке ориентировочно-исследовательской активности животных в тесте «открытое поле» нами установлено, что при профилактическом введении селенита натрия в дозе 100 мкг/кг в течение 7 дней отмечалось снижение уровня эмоциональной тревожности, о чём свидетельствует уменьшение количества стереотипных движений – груминг и болюсов. Исследовательская и двигательная активность белых крыс при ведении селенита натрия практически не изменялась. Аналогичные результаты получены в опытах с предварительным введением селенита цинка.
В ходе экспериментов с воспроизведением (УРПИ) условной реакции пассивного избегания изучаемые соединения не вызывали статистически значимое влияние на процессы обучения и памяти у интактных животных. Селенит натрия и селенит цинка не влияют на координацию движений у белых крыс.
Для характеристики действия фармакологических средств на кровообращение головного мозга при различных патологических состояниях важное значение имеют данные об их влиянии на устойчивость животных к различным видам гипоксий. Нами было изучено влияние селенита натрия и селенита цинка в дозах 30, 50, 100 мкг/кг на выживаемость животных в условиях гипоксической гипоксии. Получены результаты, согласно которым селенит натрия и селенит цинка обладают выраженным атнигипоксическим эффектом. Наибольшую активность селениты проявляли в дозе 30 мкг/кг. Так, селенит натрия повышал выживаемость животных в среднем на 22% в, а селенит цинка на 43 % сравнении с контролем.
В патогенезе органических повреждений головного мозга ведущее значение принадлежит циркуляторным расстройствам. Представляло интерес изучить влияние селенита натрия и селенита цинка на устойчивость белых крыс к циркуляторной гипоксии, вызванной билатеральной окклюзией общих сонных артерий. Селенит натрия и селенит цинка вводили внутрибрюшинно в дозах 30, 50, 100 мкг/кг за 15 минут до ишемии. На данном рисунке представлены данные о влиянии селенита натрия и селенита цинка на устойчивость белых крыс к циркуляторной гипоксии в процентном отношении к контролю. Результаты контрольной серии опытов показали, что большинство животных погибло в течение первых суток. Через 72 часа после операции в контрольной группе число выживших животных составило 37,5%. Нами установлено, что исследуемые соединения обладают антигипоксической активностью.
Селенит натрия более эффективен при его введение в дозе 50 мкг/кг (выживаемость животных к концу наблюдения составила 62,5%, что превышает контрольные значения на 25%). Аналогичный эффект проявлял селенит цинка.
Учитывая антигипоксическую активность селенитов при циркуляторной гипоксии представляло интерес изучить их действие на устойчивость организма животных к критическим гравитационным нагрузкам, краниокаудальный вектор которых вызывает нарушение кровоснабжение головного мозга, протекающее по типу ишемической гипоксии.
Центрифугирование животных проводили в 2 этапа. На первом этапе исследования крысы подвергались воздействию перегрузок величиной 16 g в течение 10 минут, после чего центрифугу останавливали на 5 минут. На втором этапе исследований, по истечении 5 минутного перерыва, животные подвергались перегрузкам величиной 21 g в течение 5 минут.
При профилактическом введение селенита натрия в дозе 100 мкг/кг на 1 этапе исследований количество выживших животных превышало контроль на 12,5%. На втором этапе на 20%.
Селенит цинка проявлял более выраженный защитный эффект: на первой стадии эксперимента не погибло ни одного животного. Выживаемость крыс составила 100%. На втором этапе исследований количество выживших животных превышало контроль 40%.
В каждой группе животных в дальнейшем проводилось тестирование психоневрологического статуса.
В течение 3 минут пребывания крысы в «открытом поле» регистрировали двигательную активность (число пересечённых секторов), ориентировочно-исследовательскую активность (вставание на задние лапы) и эмоциональный статус (количество стереотипных движений - груминг, дефекаций, уринаций, а также число посещений центрального сектора).
Исследование проводили на второй, четвёртый и седьмой день после перенесённых перегрузок. Это позволило следить за динамикой изменений психоневрологического статуса ишемизизованных животных. Полученные экспериментальные данные представлены в таблице.
В постишемическом периоде в контрольных опытах отмечалось снижение двигательной активности, о чём свидетельствует уменьшение числа пересечённых секторов, снижение познавательной активности (уменьшение числа вертикальных стоек) крыс и их эмоционального состояния (уменьшение актов груминга и числа дефекаций). Эти нарушения наиболее ярко проявлялись в течение первых дней после ишемии, а в последующие сутки несколько ослабевали. Селенит натрия и селенит цинка повышали двигательную активность крыс, перенесших ишемию мозга. Более выраженный эффект оказывал селенит цинка, под влиянием которого значительно увеличивалось число пересечённых животными секторов. Профилактическое введение селенита натрия уменьшало у крыс число вертикальных стоек, стереотипных движений и не изменяло число дефекаций по сравнению с контролем, что свидетельствует о некотором снижении исследовательской и эмоциональной активности. Селенит цинка повышал ориентировочно-исследовательскую активность (увеличивал число стоек) и снижал уровень эмоционального напряжения (уменьшал акты груминга и дефекаций) крыс в постишемическом периоде. Лучшим выражением уменьшения страха у животных является посещение ими центрального сектора, которое в наших опытах постепенно становилось более интенсивным от опыта к опыту.
При выработке УРПИ после ишемии мозга отмечалось ухудшение обучаемости крыс, что выражалось в уменьшении латентного периода захода в тёмный отсек и в увеличении длительности пребывания в нём. Селенит натрия и селенит цинка при профилактическом введении оказывают положительное влияние на процессы обучения и воспроизведения памятного следа: значительно увеличивают латентный период захода в тёмный отсек и уменьшают время пребывания в нём.
Следующим этапом исследований была оценка координации движений крыс пробой с «вращающимся стержнем». Неспособность животных удерживать равновесие на стержне в течение трёх минут рассматривали как проявление нарушения координации движений. Для изучения координации движений в динамике исследование проводили на 2, 4 и затем на 7 сутки после ишемии.
Проведённые эксперименты показали, что в постишемическом периоде в группе контрольных животных отмечалось ухудшение координации движений. Селенит цинка и селенит натрия значительно ослабляли вызванное ишемией нарушение координации движений, что проявлялось в увеличении времени удержания крыс на вращающемся стержне. Так, в контрольных опытах количество животных, удержавшихся на вращающемся стержне в течение 3-х минут, составило 42,9%. При предварительном введении селенита натрия и селенита цинка значительно повышался процент крыс, удержавшихся на стержне, и к концу недели эти показатели составили соответственно: 62,5% и 54,6% .
Таким образом, результаты анализа психоневрологического статуса в постишемическом периоде показали, что у животных наблюдается изменение всех компонентов поведения: снижение горизонтальной и вертикальной двигательной активности, дефицит эмоциональности, недостаточность мнестических функций и нарушении координации движений. Исследуемые соединения при профилактическом введении оказывают нейропротекторное действие в условиях острого нарушения мозгового кровообращения, снижают смертность животных, положительно влияют на двигательную и исследовательскую активность, уменьшают тревожность, обладают антиамнестическим эффектом, улучшают координацию движений.
При введении исследуемых соединений после ишемии мозга (лечебное действие) нами также установлена их способность устранять неврологический дефицит и улучшать высшие интегративные функции ЦНС в постишемическом периоде. Однако лечебные дозы были менее эффективными, чем профилактические.
Известно, что в постишемическом периоде ярко проявляются цереброваскулярные феномены: вначале наблюдается короткий период гиперперфузии, а затем мозговой кровоток падает значительно ниже нормы.
Особый интерес представляет вторая фаза постишемического периода. По данным В.А. Неговского, В.Г. Красильникова фаза невосстановления мозгового кровотока может усугублять повреждение мозговой ткани.
Селенит натрия существенно изменяет динамику развития постишемических цереброваскулярных феноменов, уменьшает степень проявления 1 фазы и тормозит развитие фазы невосстановления мозгового кровотока. Одинаковой по характеру и величине была реакция сосудов мозга на введение селенита цинка.
Таким образом, в сравнительном аспекте оба соединения представляют несомненный интерес в качестве средств, для коррекции церебральных ишемий – самого распространённого сейчас вида сосудистой патологии мозга. Особенности их защитного действия, вероятно, обусловлены сочетанием в спектре фармакологической активности ряда эффектов – антиоксидантного, антигипоксического, анксиолитического и ноотропного.
ОБЩИЕ ВЫВОДЫ
1. Селенит натрия и селенит цинка вызывают снижение мозгового кровотока у наркотизированных животных. С увеличением дозы соединений степень снижения кровотока уменьшается
2. Исследуемые соединения не оказывают существенного влияния на системное артериальное давление и частоту сердечных сокращений у бодрствующих животных
3. Длительное профилактическое введение селенита натрия и селенита цинка вызывает снижение уровня эмоциональной тревожности и не влияет на двигательную и исследовательскую активность, координацию движений и процессы обучения и памяти у интактных животных
4. Селенит натрия и селенит цинка обладают выраженным антигипоксическим эффектом: повышают устойчивость животных к гипоксической и циркуляторной гипоксиям
5. В условиях острой сосудистой патологии мозга селенит натрия и селенит цинка снижают смертность животных, повышают двигательную и исследовательскую активность, уменьшают тревожность, улучшают процессы обучения и памяти, координацию движений.
6. Предварительное введение селенита натрия и селенита цинка до ишемии мозга уменьшает степень проявления фазы гиперперфузии и тормозит развитие фазы невосстановления мозгового кровотока
СПИСОК ЛИТЕРАТУРЫ
1. Скворцова, В.И. Лечение и профилактика ишемического инсульта / Скорцова В.И., Стаховская Л.В. // Диагностика и терапия в клинике внутренних болезней: лекции для практикующих врачей, 10 Рос. нац. конгр. – М., 2004. - С. 142-160.
2. Кузнецов, Г.П. Клиническое значение селенодефецита у больных с сердечно-сосудистыми заболеваниями самарского региона и его коррекции препаратом «Cелена» / Г.П. Кузнецов, П.Л. Лебедев // Эксперим. и клинич. фармакология. - 1995. - Т.58, №5. С. 26-28.
3. Николаев, С.М. Влияние селенита натрия на течение экспериментального инфаркта миокарда / С.М. Николаев // 3 Всерос. съезда фармацевтов (3, 1975, Свердловск). - Свердловск, 1975. – С. 346-347.
4. Савина, М.Д. Перспектива поиска антиаритмических средств с противоишемическим эффектом, среди селенсодержащих веществ / М.Д. Савина, А.Н. Кудрин // Фармация. – 1992. Т.12, №1. - С 39 – 46.
5. Stryer, L. Biochemistry / L. Stryer - 4th ed San Francisco: Freeman, 1995. - P. 731
6. Tarp, U. Selenium in rheumatoid arthritis: a review / U.Tarp // Analist. - 1995. Vol. 120. - P. 877-881
7. Artbur, J.R. New metabolic roles for selenium / J.R. Artbur // Proc. Nutr. Soc. -1994. - Vol. 53. - P 615-624.
8. Блинова, А.М. Регуляция мозгового кровообращения / А.М. Блинова, М.Е Маршак // Физиологические механизмы регуляции мозгового кровообращения. – Л., 1963. – С. 2-41.
9. Верещагин, Н.В. Патология вертобазиллярных систем и нарушение мозгового кровообращения / Н.В. Верещагин. – М.: Медицина, 1980. – 311 с.
10. Гаевый, М.Д. Фармакология мозгового кровообращения / М.Д. Гаевый. – М.: Медицина, 1980. – 190 с.
11. Демченко, И.Т. Кровоснабжение бодрствующего мозга / И.Т. Демченко. – Л.: Наука, 1983. – 174 с.
12. Воробьева, Е.А. Анатомия и физиология: Учеб. для учащихся мед. и фармац. институтов / Воробьева Е.А., Губарь А.В., Сафьянникова Е.Б. – М.: Медицина, 1981. – 416 с.
13. Москаленко, Ю.Е. О взаимосвязи микро - и макроуровней в функциональной организации деятельности сосудистой системы головного мозга / Ю.Е. Москаленко // Физиол. журн. СССР им. Сеченова. – 1984. – Т. 70, №11. – С. 1484-1497.
14. Мотавкин, П.А. Гистофизиология сосудистых механизмов мозгового кровообращения / Мотавкин П.А., Черток В.М. – М.: Медицина, 1980. – 188 с.
15. Угрюмов, В.М. Регуляция мозгового кровообращения / Угрюмов В.М., Теплов С.И., Тиглиев Г.С. – Л.: Медицина, 1984. – 54 с.
16. Колоссовский, Б.Н. Циркуляция крови в мозгу / Б.Н. Колоссовский. – М.: Медгиз, 1951. – 372 с.
17. Мчедлишвили, Г.И. Механизмы регуляции мозгового кровообращения / Г.И. Мчедлишвили // Успехи физиол. наук. – 1980. – Т. 12, №3. – С. 75-83.
18. Физиология ЦНС: Учеб. пособие. – Ростов н/Д: Феникс, 2000. - 450 с.
19. Davis Reginald, J. Role of oxigen free radicals in focal brain ischemia / Davis Reginald J., Burcley Gregory B., Traysman Richard J. // Cerebral Hyperemia and Ischemia: From Standpoint Cerebral Blood Vol.: Proc. Sattelsump., Brain Sec. 4 th Word Congr. Microcirc., Osaca, 1-2 Aug., 1987. Amsterdam, 1988. – P. 151-155.
20. Балуева, Т.В. К вопросу о центральной норадренергической регуляции мозгового кровообращения / Т.В. Балуева // Физиол. журн. СССР им. Сеченова. – 1983. - №7. - С. 913-917.
21. Анатомия человека: В 2 т. / Под ред. М.Р. Сапина. - М.: Медицина, 1987. - Т.2. - 479 с.
22. Бородуля, А.В. Холинергическая иннервация сосудов основания головного мозга / А.В. Бородуля, Е.К. Плечкова // Журн. невропатол. и психиатрии. – 1976. - №7. С. 707-714.
23. Красильников, В.Г. Адренергические механизмы в регуляции артериальной и венозной части сосудистого русла головного мозга / В.Г. Красильников // Междунар. симпоз. по регуляции емкостных сосудов (3; 1977; Москва): тез. докл. М., 1977. – С. 74-80.
24. Куприянов, В.В. Нервный аппарат кровеносных сосудов головного мозга / Куприянов В.В., Жица В.Т. – Кишинев: Штминца, 1975. – 225 с.
25. Мирзоян, Р.С. Нейрохимический контроль мозгового кровообращения / Р.С. Мирзоян // Фармакология и токсикология. – 1983. - №4. – С. 5-15.
26. Москаленко, Ю.Е. Реактивность мозговых сосудов: физиологические основы, критерии оценки, информативная значимость / Ю.Е. Москаленко // Физиол. журн. СССР им. Сеченова. – 1986. – Т. 72, №8. – С. 1115-1122.
27. Мчедлишвили, Г.И. Физиологические механизмы регулирования микроциркуляции в коре мозга / Г.И. Мчедлишвили, Д.Г. Бералидзе // Физиол. журн. СССР им. Сеченова. – 1984. – Т. 70, №11. – С. 1476-1483.
28. Ташаев, Ш.С. Об адаптивной функции симпатической иннервации мозговых сосудов при быстрых изменениях системного артериального давления / Ш.С. Ташаев // Бюл. экспирим. биологии и медицины. – 1980. – Т. 90, №11. – С. 543-546.
29. Siesje, B. K. Mechanisms of ischemic damage to neurons, glial cells and vascular tissue / B. K. Siesje, Maj-Lie. Smith // Regul. Mech. Neuron. Vessel Commun. Brain: Prog. NATO Adv. Res. Workshop, Salo, sepr. 3-8, 1988. – Berlin, 1989. – P. 209-223.
30. Сравнительная характеристика некоторых гемодинамических и биохимических показателей постишемического периода в зависимости от режимов аутогемоперфузии мозга / М.Д Гаевый [и др.] // Физиол. журн. СССР им. Сеченова. – 1991. - №77. – С. 76-80.
31. Burnstoer, X. Comprative studies of purinergic nerves / X. Burnstoer // J. Exp. Zool. – 1975. – Vol. 194, № 1. – P. 103-133.
32. Демченко, И.Т. Метаболические факторы регуляции / И.Т. Демченко // Физиология кровообращения. Регуляция кровообращения. – Л.: Наука, 1986. – С. 167-193.
33. Faraci, F.M. Myogenic mechanisms in the cerebral circulation / F.M. Faraci, G.L. Bumbach, D.D. Heisad // J. Hypertens. – 1989. – Vol. 7, № 4. – P. 61-64.
34. Mechanisms of drug-induced vasodilatation / Gurney Alison M. [et al.] // J. Pharm. and Pharmacol. – 1994. – Vol. 46, № 4. – P. 242-251.
35. Москаленко, Ю.Е. Принципы изучения сосудистой системы головного мозга человека / Москаленко Ю.Е., Хилько В.А. – Л.: Наука, 1984. – 68 с.
36. Москаленко, Ю.Е. Мозговое кровообращение: физико-химические приемы изучения / Москаленко Ю.Е., Бекетов А.И., Орлов Р.С. – Л.: Наука, 1988. – 159 с.
37. Mc. Calden, T. The role of the carotid chemoreceptors in the cerebrovascular hypercapnic dilatation / T. Mc. Calden, C. Rosendorff // Acta Neurol. Scand. – 1977. – Vol. 56, № 64. – P. 292-293.
38. Бархатова, В.П. Нейротрансмиттерная организация двигательных систем головного и спинного мозга в норме и патологии / В.П. Бархатова, И.А. Завалишин // Журн. неврологии и психиатрии. - 2004. - №8. - С. 77-82.
39. Кулинский, В.И. Предача и трансдукция гормонального сигнала в разные части клетки / В.И. Кулинский // Соровский образовательный журн. – 1997. - №8. - С. 14-19.
40. Ткачук, В.А. Молекулярные механизмы нейроэндокринной регуляции / В.А. Ткачук // Соровский образовательный журн. – 1998. - №6. - С. 16-20.
41. Скулачев, В.П. Явления запрограммированной смерти. Митохондрии, клетки и органы: роль активных форм кислорода / В.П. Скулачев // Соровский образовательный журн. – 2001. – Т. 7, №6. – С. 4-10.
42. Ковалев, Г.В. Поиск веществ, активирующих ГАМК-ергическую систему, новое направление в создании антигипертензивных средств / Г.В. Ковалев, И.Н. Тюренков // Фармакология и токсикология. - 1989. - №1. - С. 5-11.
43. Тюренков, И.Н. Роль гамкергической системы мозга в регуляции кровообращения / И.Н. Тюренков, В.И. Перфилова // Эксперим. и клинич. фармакология. – 2001. - Т. 664, 6. - С. 68-72.
44. Baranano, D.E. Atipical neural messengers / D.E. Baranano, C.D. Ferris, S.H. Snyder // Treds Neurosci. - 2001. – Vol. 24. - P. 99-106.
45. Bunin, M.A. Paracrine neurotransmission in the CNS: involment of 5-HT / M.A. Bunin, R.M. Wightman // Treds Neurosci. - 1999. – Vol. 22. - P. 377-382.
46. Кулинский, В.И. Нейротрансмиттеры и головной мозг / В.И. Кулинский // Соросовский образовательный журн. – 2001. – Т. 7, №6. – С. 11-16.
47. Саратиков, А.С. Экспериментальная и клиническая фармакология мозгового кровообращения / Саратиков А.С., Белоспасов В.В., Плотников М.В. – Томск: Изд-во Томс. ун-та, 1979. – 248 с.
48. Сихарулидзе, Н.В. Роль ацидоза в развитии постишемического отека мозга у кроликов / Н.В. Сихарулидзе, Г.И. Мчедлишвили // Патол. физиология и эксперим. терапия. – 1987. - №3. – С. 47-49.
49. Alberti, K. The hydrogen ion normal metabolism: a review // Metabolic acidosis. (Ciba found. symposium 87) / Alberti K., Cuthbert C. London, 1982. – P. 1-19.
50. Meyer, J.S. Central neurogenic control of cerebral blood flow / J.S. Meyer, T.Teraura, K. Secamoto // Neurology. – 1971. – Vol. 21. – P. 247-262.
51. Мирзоян, Р.С. Нейрохимический контроль мозгового кровообращения / Р.С. Мирзоян // Фармакология и токсикология. – 1983. - №4. – С. 5-15
52. Кораблев, М.В. Противогипоксические средства / Кораблев М.В., Лукиенко П.И. Минск: Беларусь, 1976. – 128 с.
53. Плотников, М.В. Антиокислительные эффекты антигипоксантов при ишемии мозга / М.В. Плотников, Е.А. Кобзева, Т.М. Плотникова // Бюл. экспирим. биологии и медицины. – 1992. - №5. – С. 504-506.
54. Потапов, А.А. Результаты масспектрометрических исследований напряжения кислорода, углекислого газа и кровотока в ткани мозга у нейрохирургических больных / Потапов А.А., Сировский Э.Б., Маневич А.З. // Вопр. нейрохирургии. 1979. - №1. - С. 20-26.
55. Салалыкин, В.И. Гипоксия головного мозга (клиника и лечение) / Салалыкин В.И., Арутюнов В.В. – М.: Медицина, 1978. – 196 с.
56. Mellander, S. Myogenic mechanisms in local vascular control / S. Mellander // Int. Congr. Physiol. Sci. “31, 1989, Helsinki”. – Oulu, 1989. – P. 7-8.
57. Березовский, В.А. Гипоксия и индивидуальные особенности организма / В.А. Березовский – Киев: Наук. думка, 1978. – 279 с.
58. Петров, И.Р. Кислородное голодание мозга (экспериментальные материалы) / И.Р. Петров – Л.: Медгиз, 1949. – 48 с.
59. Динамика развития постишмических цереброваскулярных феноменов в условия перфузии мозговых сосудов стабильным объемом крови / М.Д. Гаевый [и др.] // Патол. физиология и эксперим. терапия. – 1982. – Вып. 4. – С. 61-64.
60. Орлов, Р.С. Механизмы расслабления гладких мышц сосудов / Р.С. Орлов // Актуальные вопросы физиологии кровообращения. – Симферополь, 1980. – С. 117-123.
61. Evans, M.C. The effect of local changes in calcium concentration on hypotalamic blood flow in the anastesiel rabbit / M.C. Evans, R.A. Linton, I.G. Cameron // Cereb. blood flow and metabolism. – Munskgaard, 1977. – P. 388- 389.
62. Weibenger, J. Cerebral blood flow in the evolution of infarction following unilateral carotid artery oclusion in mongolian gerbils / J. Weibenger, Nieves-Rosa J. // Stroke. – 1987. – Vol. 18, № 3. – P. 612-615.
63. Митагвария, Н.П. Функциональное значение отдельных звеньев механизма регуляции мозгового кровообращения / Н.П. Митагвария // Физиолология, патофизиология и фармакология мозгового кровообращения: тез. докл. 2 Всесоюз. конф. 10-12 нояб., 1988 г. – Тбилиси, 1988. – С. 123.
64. Мотавкин, П.А. Гистофизиология сосудистых механизмов мозгового кровообращения / Мотавкин П.А., Черток В.М. – М.: Медицина, 1980. – 188 с.
65. Напряжение кислорода и локальный кровоток в мозгу животных при изменении состава дыхательной смеси / А.И. Селивра [и др.] // Физиология, патофизиология и фармакология мозгового кровообращения.: тез. докл. 2 Всесоюз. конф. 10-12 нояб., 1988 г. – Тбилиси, 1988. – С. 165.
66. Сравнительная характеристика некоторых гемодинамических и биохимических показателей постишемического периода в зависимости от режимов аутогемоперфузии мозга / М.Д. Гаевый [и др.] // Физиол. журн. СССР им. Сеченова. – 1991. - №77. С. 76-80.
67. Иванов, К.П. Успехи и спорные вопросы в изучении микроциркуляции / К.П. Иванов // Физиол. журн. СССР им. Сеченова. – 1995. – Т. 81, №6. – С. 1-21.
68. Теплов, С.И. Гормональные факторы регуляции кровообращения / С.И. Теплов // Физиология кровообращения – Л., 1986. – С. 94-110.
69. Неретин, В.Я. Гормональная регуляция мозгового кровотока / В.Я Неретин, С.В. Котов, Г.С. Молчанова // Журн. невропатол. и психиатрии. – 1990. – Т. 90, №7. С. 41-43.
70. Азин, А.Л. Аминергический контроль артерий головного мозга / А.Л. Азин // Успехи физиол. наук – 1983. – Т. 14, №2. – С. 98-116.
71. Балуева, Т.В. О симпатическом тонусе мозговых сосудов / Т.В. Балуева, Н.И. Гирс, С.И. Теплов // Физиол. журн. СССР им. Сеченова. – 1982. - №5. – С. 596-600.
72. Lee, T. Direct evidence against acetylcholine as the dilatator transmitter in the cat cerebral artery / T. Lee // Europ. J. Parmacol. – 1980. – Vol. 58 – P. 393-397.
73. Mаtsuda, M. Effect of acetylcholine on cerebral circulation / M. Mаtsuda, J. Meyer, V. Deshmukh // J. Neurosung. – 1976. – Vol. 45. – P. 423-431.
74. Митагвария, Н.П. Нейрогенный контроль регуляции мозгового кровообращения и сопряженность кровоток-метаболизм-функция / Н.П. Митагвария // Физиол. журн. СССР им. Сеченова. – 1989. – Т. 75, №11. – С. 1473-1480.
75. Dopaminergic action on brain vessels. / Z. Edvinsson [et al.] // Intern. Catecholamine sump. (4; 1979, New York): abstr…. – New York, 1979. – P. 1494-1496.
76. Edvinsson, Z. Neurogenic mechanisms in the cerebrovascular bed; Autonomic nerves, amine receptors and their effects on cerebral blood flow / Z. Edvinsson // Acta physiol. Scand. – 1975. – Vol. 96, Suppl. 427. – P. 1-35.
77. Meuller, S. Effect of sympatetic nerves on cerebral vessels during seizures / S. Meuller, D.D. Heistand, M. Marcus // Amer. J. Physiolog. 1979. – Vol. 237, № 2. – P. H178-H184.
78. Purves, M.J. The physiology of cerebral circulation / M.J. Purves Cambridge, 1972. – 252 p.
79. Karninski, L.P. Potential role of Cl-HCO3 exchange in regulating vascular tone / L.P. Karninski // Hypertension. – 1989. – Vol. 15, 15 – P. 104-106.
80. Логинов А.В. Физиология с основами анатомии человека: Учебник для фарм. институтов / А.В. Логинов – М.: Медицина, 1983.- 495 с.
81. Селен в организме человека / В.А. Тутельян [и др.] – М.: РАМН, 2002. - 224 с.
82. Сидельникова, В.Д. Геохимия селена в биосфере / В.Д. Сидельникова // Проблемы биогеохимии и геохимической экологии. М..: Наука, 1999. - Т. 23.- С. 81-99.
83. Ермаков, В.В. Биологическое значение селена / Ермаков В.В., Ковальский В.В. - М.: Наука, 1974. – 58 с.
84. Ермаков, В.В. Геохимическая экология как следствие системного изучения биосферы / В.В. Ермаков // Проблемы биогеохимии и геохимической экологии. - М.: Наука, 1999. - Т. 23. - С 152-182.
85. Alftban, G. The effects of selenium fertilization on glutathione peroxidase and selenoprotein P I Finland / G. Alftban // Proc. 7th Nordic Symp. On trace elements in human health and disease. Espoo, 1999. - P. 39.
86. Robinson, M.F. The moonstone: more about selenium / M.F. Robinson // J. Hum Nutr. - 1976. - Vol. 30. - P. 79-91.
87. Artbur, J.R. Roles of selenium in type I iodithyronine 5'-deiodinase and in thyroid hormone and iodine metabolism / Artbur J.R. Becrett G.J. // Selenium in biology and human health / Ed. R.F.Burk. - New York: Springer-Verlag, 1994. - P. 93-115.
88. Reiter, R. Selenium and drug metabolism. I. Multiple modulations of mous liver enzymes / R. Reiter, A. Wendel // Biochem. Pharmacol. - 1983. - Vol. 32. - P. 3063-3067.
89. Crystal structure of formate dehydrogenase H: Catalysis involving Mo, molybdopterin, selenocysteine and an Fe4S4 cluster / J.C. Boyington [et al.] // Science - 1997. - Vol. 275. - P. 1305-1308.
90. Molecular biology of selenium and its role in human health / B.J. Lee [et al.] // Mol. Cells. - 1996. - Vol. 6. - P. 509-520.
91. Testosterone mediates expression of the selenoprotein PHGPx by induction of spermatogenesis and not by direct transcriptional gene activation / M. Maiorino [et al.] // FASEB J. - 1998. - Vol. 12. - P. 1359-1370.
92. Beck, C. The selen-mediated de-ionisation of iodophenenols: A selenium model for the mechanism of 5'-thyronine de-iodinase / C. Beck, S.B. Jensen, J. Reglinski // Thyroid. - 1994. - Vol. 4. - P. 1353-1356.
93. Gladysbev, V.N. DL-selenoprotein containg proteins in mammals / V.N. Gladysbev // J. Biomed. Sci. - 1999. - Vol. 6, N 3. - P. 151-166.
94. Levander, O. Selenium / O. Levander, R.F. Burk // Present knowledge in nutrition / Eds. E.E. Ziegler, L.J. Filer. N.Y.: Acad. Press, 1998. - Vol. 20. - P. 4492-4500.
95. Selenocysteine lyase, a novel enzyme that specifically acts on selenocysteine: mammalian distribution and purification and properties of pig liver enzyme / N. Esaki [et al.] // J. Biol. Chem. - 1982. - Vol. 257. - P. 4386-4391.
96. Gantber, H.E. Metabolism of hydrogen selenide and methylated serenades / H.E. Gantber // Advanced in nutritional research / Ed. H.H. Draper. New Yeark: Plenium, 1979. - Vol. 2. - P. 107-128.
97. Mozier, N.M. S-adenosil-L-methionine: tioether S-methiltransferase a new enzyme in sulfur and selenium metabolism / N.M. Mozier, K.P. McConnell, J.W. Hoffman // J. Bol. Chem. - 1988. - Vol.263. - P. 4527-4531.
98. Bopp, B.A. Metabolitic rate of selected selenium compounds in laboratory animal and man / B.A. Bopp, R.C. Sonders, J.W. Kesterson // Drug Metab. Rev. - 1982. - Vol. 13. - P. 271-318.
99. Berri, M.J. Cloning and expression of the human selenium donorprotein, the homolog of prokaryotic / M.J. Berri, [et al.] // FASEB J. - 1995. - Vol. 9. - P. 286
100. Salbe, A.D. Selenium content of rat hair nails and other tissues is affected by concurrent exposure to toxic elements / A.D. Salbe, V.C. Morris, O.A. Levander // Nutr. Res. - 1993. - Vol. 13. - P. 31-36.
101. Human [74Se] selenomethionine metabolism: a kinetic model / C.A. Swanson [et al.] // Am. J. Clin. Nutr. - 1991. - Vol. 54. - P. 917-926.
102. Evidence for intestinal release of absorbed selenium in a form with hepatic extraction / T. Kato [et al.] // Am. J. Physiol. - 1992. -Vol. 262. - P. G.854-G858.
103. Gantber, H.E. Selenotrisulfides. Formation by the reaction of thiols with selenious acid / H.E. Gantber // Biochemistry. - 1968. - Vol. 7. - P. 2898.
104. Direct measurement of selenium absortion in vivo: triple-lumen gut perfusion in the concious dog / P.G. Reasbeck [et al.] // Proc. New Zealand Workshop on Trace Elements Dunedin, 1981. - P. 107.
105. m-RNA stability and selenocysteine insertion sequence efficiency rank gastrointestinal glutathione peroxidase high in the hierarchy of selenoproteins / K. Wingler [et al.] // Eur. J. Biochem. - 1999. - Vol. 259, N 1-2. - P. 149-157.
106. Combs, G.F. Selenium in nutrition / G.F. Combs / Encyclopedia of human biology. 2nd ed. New Yeark: Acad. Press. - 1997. - Vol. 7. - P. 743-754.
107. Studies in the distribution and characteristics of new mammalian selenium-containing proteins / D. Bebne [et al.] // Analyst. - 1995. - Vol. 120. - P. 823-825.
108. Thomson, C.D. Selenium speciation in human body fluids / C.D. Thomson // Ibid. - 1998. - Vol. 123. - P. 827-831.
109. Adsorption of selenite and selenomethionine from ligatid digestive tract segments in rats / P.D. Wbanger [et al.] // Proc. Soc. Exp. Biol. Med. - 1976. -Vol. 153. - P. 295.
110. Burk, R.F. Selenium in nutrition / R.F. Burk // World Rev. Nutr. Diet. - 1978. - Vol. 30. - P. 88-106.
111. Studies of safe maximal daily dietary selenium intake in a seleniferous area in China. I. Selenium intake and tissue selenium levels of the inhabitants / G. Yang [et al.] // J. Trace Elem. Electrolytes Health Dis. - 1989. - Vol. 3. - P. 77-87.
112. Use of selenium concentration in whole blood, serum, toenails or urine as a surrogate measure of selenium intake / M.P. Lonnecker [et al.] // Epidmiology. - 1996. - Vol. 7. - P. 384-390.
113. Селенобогащённые дрожжи Saccaromyces cerevisiae / Н.А. Голубкина [и др.]. // Биотехнология. - 1996. - № 75. - С. 52-57.
114. Effects of selenium supplementation for cancer prevention in patients with carcinoma of the skin / L.C. Clark [et al.] // JAMA. - 1996.- Vol. 276, N 26. - P. 1957-1963.
115. Rosenfeld, I. Selenium: geobotany, biochemistry, toxicity and nutrition / Rosenfeld I., Beath O.A. - N.Y.: Acad. Press, 1964.
116. Studies of safe maximal daily dietary Se- intake in a seleniferous area in China. II. Relation between Se- intake and the manifestation of clinical signs and certain biochemical alterations in blood and urine / G. Yang [et al.] // Ibid. – 1976. - P. 123-130.
117. Yang, G. Human selenium requirements in China / G. Yang, L. Zbu, S. Liu // Selenium in biology and medicine. - Westport: AVI, 1987. - P. 589-607.
118. Studies on the relations of selenium and Keshan disease / X. Cben [et al.] // Biol. Trace Elem. Res. - 1980. - Vol. 2. - P. 91-107.
119. Burk, R.F. Selenium in nutrition / R.F. Burk // World Rev. Nutr. Diet. -1978. - Vol. 30. - P. 88-106.
120. Jobnson, R.A. An accidental case of cardiomyopathy and selenium deficiency / R.A. Jobnson, S.A. Bker, J.T. Fallon // N. Engl. J. Med. - 1981. -Vol. 304. - P. 1210-1212.
121. Association between cardiovascular death and myocardial infarction and serum selenium in a matched-pair longitudinal study / J.T. Salonen [et al.] // Lancet. - 1982. - Vol. 2. - P. 175-179.
122. Changes in smoking, serum cholosterol and blood pressure levels during a community-based cardiovascular death prevention program - the North Karelia Project / J.T. Salonen [et al.] // Am.J. Epidemiol. - 1981. - Vol. 114. - P. 81-94.
123. Iodine and selenium deficiency associated with cretinism in northern Zaire / J.B.Vanderplas [et al.] // Am. J. Clin. Nutr. - 1990. - Vol. 52. - P. 1087-1093.
124. Selenium dependent and selenium non-dependnt GSHPx in the patients suffering from Balkan endemic nephropathy / G. Grubo-Lajsic [et al.] // Proc. Int. Symp. «Selenium in geochemistry, biology and medicine». Belgrad, 1996. - P. 69.
125. Moxon, A.L. Selenium poisoning / A.L. Moxon, M.A. Rbian // Physiol. Rev. - 1943. - Vol. 23. - P. 305.
126. Palmer, I.S. Toxicity of various selenium derivatives to chick embryos / I.S. Palmer, R.L. Arnold, C.W. Carlson // Poultry Sci. - 1973. - Vol. 52. - P. 1841-1844.
127. Levander, O.A. Dietary selenium levels needed to maintain balance in North American adults consuming self-selected diets / O.A. Levander, V.C. Morris // Am. J. Nutr. - 1984. - Vol. 39. - P. 809-815.
128. Essentiality versus toxicity some considerations in the risk assessment of essential trace elements / C.O. Abernatby [et al.] // Hazard assessment of chemicals / Ed. J. Saxena. Vol. 8. Washington: Taylor and Francis, 1993. - P. 81-113.
129. О радиозащитных свойствах селенсемикарбазида / Г.Б. Абдуллаев [и др.] // Селен в биологии. – Баку, 1976. – 236 с.
130. Якубенко, А.В. Лучевая болезнь / А.В. Якубенко – Л., 1966. – 269 с.
131. Голубкина, Н.А. Селен в сыровотки крови и опухоли у больных с доброкачественными и злокачественными новообразованиями / Н.А. Голубкина, М.Д. Алиев, Н.Е. Кушлинский // Вопросы мед. химии. – 1995. – Т. 41, №4. – С. 50 – 54.
132. Противоопухолевая активность производных селенабициклонида / Б.М. Бутин [и др.] // Хим.-фармац. журн. – 1995. – Т. 29, №8. – С. 18-19.
133. Синтез и радиозащитный эффект производных тиазиазолов, тиадизолинов и их селеновых аналогов / Л.П. Петров [и др.] // Хим.-фармац. журн. – 1994. – Т. 28. №2 – С. 19-22.
134. Taylor E.W. Selenium and cellular immunity // Biol. Trace. Res. - 1995. - Vol. 49. - P. 85-89.
135. Porter J.M. Antioxidant therapy in the prevention of organ dysfunction syndrome and ihfection complications after trauma: early results of a prospective randomized study // Am. Surg. - 1999. - Vol. 65, N 5. - P. 478-483.
136. О возможности регуляции свободнорадикального состояния сетчатки глаза соединениями селена / Г.Б. Абдуллаев [и др.] // Селен в биологии. – Баку, 1974. - 159 с.
137. Абдуллаев, Г.Б. Изучение влияния селенсодержащих пищевых экстрактов на зрительную функцию при дистрофиях сетчатки / Г.Б. Абдуллаев [и др.] // Селен в биологии. – Баку, 1976. – 250 c.
138. Наздрюха, Л.Р. Биологическая роль микроэлементов в организме животных и человека / Л.Р. Наздрюха – М.: Наука. – 1977. – 184с.
139. Кудрин, А.Н. Фармакотерапия препаратами селена экспериментального гепатита / Кудрин А.Н., Левшин Б.И., Мехтиев М.А. – Баку, 1982. – 221 с.
140. Венцславская, Т.А. Противоаритмическая активность нового селенопроизводного препарата / Т.А. Венцславская, Л.Л. Стажадзе, В.В. Коржова // Фармакология и токсикология. - 1984. - №2. – С. 38 – 41.
141. Selenium glutathione peroxidase, and platelet functions / D. Vitoux [et al.] // Ann. Biol. Clin. - 1996.-Vol. 54. - P. 181-187.
142. The enzyme glutathione peroxidase in arachidonic acid metabolism of human platelets / G. Guidi [et al.] // J. Lab. Clin. Med. - 1984. - Vol. 104. - P. 574-577.
143. Vader, C.R. Pulmonary prostaglandin metabolism during normobaric hyperoxia / C.R. Vader, M.M. Matbias, C.L. Scbatte // Prostagland. Med. - 1981. - Vol. 6. - P. 101-105.
144. Effect of sodium selenite on in vitro interactions between platelets and endothelial cells / M.M. Ricetti [et al.] // Int. J. Clin. Lab. Res. - 1999. - Vol. 29, N 2. - P. 80-84.
145. Левшин, Б.И. Влияние препаратов селена на некоторые показатели белкового жирового и углеводного обмена у животных с токсическим гепатитом / Б.И. Левшин // Актуальные вопросы фармации Вып. 2: Сб. науч. тр. – Ставрополь, 1974. – С. 363 364.
146. Левшин, Б.И. Новые гепатопртекторные вещества из препаратов селена / Б.И. Левшин // Актуальные проблемы фармакологии и фармации: Сб. науч. тр. – М., 1971. – С. 90 97.
147. Левшин, Б.И. Экспериментальная фармакотерапия препаратами селена и тазолидина токсического повреждения печени: Афтореф. дис. … д-ра. мед. наук / Б.И. Левшин Харьков, 1973. – 43 с.
148. Selenium in acute infection / K. Sammalkorpi [et al.] // Infection. - 1988. -Vol. 16, N 4. - P. 222-224.
149. Шарипов, К.О. Роль органических производных селена в регуляции антиокислительных процессов в печени при экспериментальном токсическом гепатите / К.О. Шарипов // Вопр. биолог. мед. и фармац. химии. - 2002. - №3. - С. 40 – 44.
150. Модуляция инфекционного процесса препаратами бромокриптина, соматотропина и селена у мышей, аналогичная связь между тяжестью инфекционного заболевания (вирусный гепатит) и уровнями селена, соматотропина, пролактина в крови больных людей / Н.А. Голубкина [и др.] // Иммунология. - 1997. - №6. - С. 27-29.
151. Эффект обогащения рационов селеном и витамином Е на некоторые показатели иммунитета у крыс / Э.Н. Трушина [и др.] // Бюл. эксперим. биологии и медицины. - 1995. – Т. 11. №3 – С. 317-319.
152. Бернштейн, Т.Ф. Влияние молибдена и селена на некоторые показатели естественного иммунитета у кроликов / Т.Ф. Бернштейн, Т.А. Кардович, З.С. Клеванович / Сб. научн. трудов Витебского мед. ин-та, вып. 13, кн. 2. – Витебск, 1969. – С. 285 – 289.
153. Selenium in the testis of the rat: studies on its regulation and its importance for the organism / D. Bebne [et al.] // Ibid. - 1982. - Vol. 112. - P. 1682-1686.
154. Effect of selenium, vitamin E end antioxidants on testicular function in rats / A.S. Wu [et al.] // Biol. Reprod. – 1973. – Vol. 8. P. 625-629.
155. Нежданов, А.Г. Влияние селенита натрия на стероидогенез у животных / А.Г. Нежданов, С.А. Власов // Тез. докл.: науч. конф. «Микроэлементы в биологии и их применение в сельском хозяйстве и медицине». – Самарканд, СамГУ, 1990. - С. 374-375.
156. Экологические проблемы нарушения сперматогенеза у спортсменов и молодёжи / Р.Д. Сейфула [и др.] // Сб. науч. тр. ВНИФК. - М., 1997. – С. 322-333.
157. Levander, O.A. Scientific rational for the 1989 recommended dietary allowance for selenium / O.A. Levander // J. Am. Diet Ass. - 1989 - Vol. 91, N. 12. - P. 1572-1576.
158. Djujic, I.S. Redistribution of selenium among tissues effected by radiation / I.S. Djujic, O. Jozanov-Stankov // Biol. Trace Elem. Res. - 1992. - Vol. 33. - P. 197-204.
159. Кулагин, О.Л. Гепатопротекторное действие карсила, масла расторопши пятнистой, препарата «Селена» при токсическом гепатите / О.Л. Кулагин, Е.В. Кантория // Разработка, исследование и маркетинг новой фармацевтической продукции: Сб. науч. тр. ПятГФА. Вып.60. – Пятигорск, - 2005. С. 364 - 366.
160. Kralick, F.A. Method for the inhibition of the proliferation of cancer cells in a tumorsensitive to treatment with a selenodithiol by the injecthiol into the tumor of a selenodithiol such as selenodiglutathione / F.A. Kralick, W.B. Parrisb, D.N.Willett // USA Patent. - 1992. - N 5. - P. 852.
161. Carcinogen-induced chromosomal breakage decreased by antioxidants / R.J. Sbamberger [et al.] // Proc. Natl. Acad. Sci. USA. - 1973. - Vol. 70. - P. 1461-1463.
162. Rosin, M.P. Assessment of the use of the Salmonela mutagenesis assay to determine the influence of antioxidants on carcinogen-induced mutagenesis / M.P. Rosin, H.F. Sticb // Int. J. Cancer. - 1979. - Vol. 23. - P. 722-727.
163. Sbamberger, R.J. Antioidats reduce he mutagenic effectof malonaldehyde and βpropiolacone. Partix, antioxidas and cancer / R.J. Sbamberger [et al.] // Mutat. Res. - 1979. - Vol. 66. - P. 349-355.
164. Rosin, M.P. Inhibition of spontaneous mutagenesis in yeast cultures b selenite, selenate and selenide / M.P. Rosin // Cancer Lett. - 1981. - Vol. 13. - P. 7-14.
165. Lawson, T. Enhancement of the repain of carcinogen-inducted DNA damage in the human lymphocytes / T. Lawson, D.F. Birt // Hereditas. - 1983. -Vol. 98. - P. 249-252.
166. Combs, G. The role of selenium in nutrition / Combs G., Combs S. - New Yeark.: Acad. Press, 1986. – P. 123.
167. Clayton C.C. Diet and azo dye tumors: effect of diet during a period when the drug is not fed / C.C. Clayton, C.A. Baumann // Cancer Res. - 1949. - Vol.9. - P. 575.
168. Scbrauser, G.N. Cancer mortality correlation studies. IV. Associations with dietary intakes and blood levels of certain trace elements, notable Se-antagonists / G.N. Scbrauser, D.A. Wbite, C.J. Scbneider // Bioiorg. Chem. - 1977. - Vol. 7. -P. 35.
169. Scbrauser, G.N. Cancer mortality correlation studies. III. Statistical association with dietary selenium intakes / G.N. Scbrauser, D.A. Wbite, C.J. Scbneider // Bioiorg. Chem. - 1977. - Vol. 7. - P. 23.
170. Sbamberger, R.J. Selenium distribution of human cancer mortality / R.J. Sbamberger, C.E. Willis // Crit. Rev. Clin. Lab. Sci. - 1971. - Vol. 2. - P. 211.
171. Willett, W. Prediagnostic serum selenium and risk of cancer / W. Willett, B. Polk, S. Morris // Lancet. - 1983. - Vol. 2. - P. 130-134.
172. Risk of cancer in relation to serum concentrations of selenium and vitamins Amand E / J.T. Salonen [et. al.] // Br. Med. J. - 1985. - Vol. 290. - P. 417-420.
173. Plasma selenium and skin neoplasma / L.C. Clark [et al.] // Nutr. Cancer. -1984. - Vol. 7. - P. 743-754.
174. Effect on an aqueous extract of selenium-enriched garlic on in vitro markers and in vivo efficacy in cancer prevention / J. Lu [et al.] // Carcinogenesis. - 1996. - Vol. 17. - P. 1903-1907.
175. Влияние b-каротина на усвоение селена в норме и патологии / Голубкина М.А. [и др.] // «Питание и здоровье. Биологически активные добавки к пище». Тез. докл: Междунар. Симпоз. - М., 1996. - С. 36.
176. Кудрявцева, Л.А. Влияние селенита натрия и витамина Е на животных: Афтореф. дис. … д-ра. мед. наук / Л.А. Кудрявцева – М., 1967. – 21 с.
177. Welsb, S.O. The protective effect of vitamin E and selenium against methylmercury toxicity in the Japanese quail / S.O. Welsb, J.H. Soares // Nutr. Rep. Int. - 1976. - Vol. 13. - P. 43.
178. Влияние селена на ферменты метаболизма ксенобиотиков печени крыс / Л.В. Кравченко [и др.] // Вопр. мед. химии. – 1991. – Т.37, вып. 5. – С. 73 – 76.
179. Влияние селена на образование канцерогенных N- нитрозоминов / В.П. Дерягина [и др.] // Вопр. питания. - 1996. - №3. - С. 31-33.
180. Ингибируещее действие селена на эндогенный синтез N- нитрозосоединений / С.А. Хотимиченко [и др.] // Вопр. питания. - 1997. - №4. - С. 16-18.
181. Влияние селена на эндогенный синтез нитрозоаминов и токсическое действие нитритов у крыс / Е.М. Мамаева [и др.] // Вопр. питания. - 1994. - №4. - С. 32-34.
182. Burcк, R.F. Ethane production and liver necrosis in rats afther administration of drags and other chemicals / R.F. Burcк, J.M. Lane // Toxicol. Appl. Pharmacol. - 1979. - Vol. 50. - P. 467.
183. Parizek, J. The protective effect of smoll amounts of selenite in sublimate intoxication / J. Parizek, I. Ostadalova // Separatum Experientia. - 1967. - Vol. 23. - P. 143.
184. Selenium relation to decreased toxicity of methylmercury added to diets containing tuna / H.E. Gantber [et al.] // Science. - 1972. - Vol. 23. - P. 1122.
185. Sell, J.L. Influence of selenium on toxicity and metabolism of methylmercury in chicks and quail / J.L. Sell, F.G. Horani // Ibid.- 1987 - Vol. 14. - P. 439.
186. El-Begearmi, M.M. A mutual protective effect of mercury and selenium in Japanese quail / M.M. El-Begearmi, M.L. Sunde, H.E. Ganter // Poultry Sci.- 1977. - Vol. 56. - P. 313.
187. Holmberg, R.E. Interrelationships of selenium, cadmium and arsenic in mammalian teratogenesis / R.E. Holmberg, V.H. Ferm // Arch. Environ. Health.- 1969. - Vol. 18. - P. 873.
188. Moxon, A.L. The effect of arsenic on the toxicity of seleniferous grains / A.L. Moxon // Science. - 1938. - Vol. 88. - P. 81.
189. Cban, S. The role of copper, molybdenum, selenium and zink in nutrition and health / S. Cban, B. Gerson, S. Subramaniam // Clin. Lab. Med. - 1998. -Vol. 18, N 4. - P. 673-685.
190. Amer, M.A. Supplemental copper and selenium for calves: effects upon ceruloplasmin activity and liver copper concentration / M.A. Amer, G.J. St. Laurent, G.J. Brisson // Can. J. Physiol. Pharmacol. - 1973. - Vol. 51. - P. 649.
191. Исследование антиканцерогенных свойств микроэлемента селена в санитарно-гигиеническом эксперименте / В.А. Книжников [и др.] // Гиг. и сан. - 1993. - №7. - С.54-57
192. Руководство по экспериментальному (доклиническому) изучению новых фармакологических веществ / Под ред. В.П. Фисенко [и др.] - М.: ИАА «Ремедиум», 2000. - С. 220 - 224.
193. Измеров, Н.Ф. Параметры токсикометрии промышленных ядов при однократном воздействии: Справочник / Измеров Н.Ф., Саноцкий И.В., Сидоров К.К. - Л.: Медицина, 1977. - С. 196 - 197.
194. Демченко, И.Т. Измерение органного кровотока с помощью водородного клиренса / И.Т. Демченко // Физиол. журн. -1981. - Т.67, №1. -С. 178-183.
195. Гаевый, М.Д. Физиология, патофизиология и фармакология мозгового кровообращения / М.Д. Гаевый. - Ереван, 1984. - С. 40-41.
196. Ивашев, М.Н. Изучение центральной гемодинамики с использованием компьютерной программы «BEAT» / М.Н. Ивашев, Т.А. Лысенко, В.А. Коршунов // Человек и лекарство: Тез. докл. 4 Рос. нац. конгр. 2-6 апр. 1997г. - М, 1997. - С. 263.
197. Многократные измерения параметров системной гемодинамики у бодрствующих крыс / О.С. Медведевх [и др.] // Физиол. журн. СССР. - 1986. - Т.72, №2. - С. 253-256.
198. Маркель, А.Л. К оценке основных характеристик поведения крыс в тесте “открытое поле” / А.Л. Маркель // Журн. высш. нервн. деятельности. - 1981. - Т.31, вып. 2. С.433-443.
199. Маркель А.Л. Метод комплексной регистрации поведенческих и вегетативных реакций у крыс при проведении теста “открытое поле” / А.Л. Маркель, Р.А. Хусаинов // Журн. высш. нервн. деятельности. - 1976. - Т.31, вып. 6. – С.345-353.
200. Маркель, А.Л. Факторный анализ поведения крыс в тесте открытого поля / А.Л. Маркель, Ю.К. Галактионов, В.М. Ефимов // Журн. высш. нервн. деятельности им. Павлова. - 1988. - Т.33, вып.5. - С. 855-863.
201. Сернов, Л.Н. Элементы экспериментальной фармакологии / Сернов Л.Н., Гацура В.В. - М., 2000. - 352 с.
202. Гацура, В.В. Методы первичного фармакологического исследования биологически активных веществ / В.В. Гацура - М.: Иностр. лит., 1974.- 142 С.
203. Буреш, Я Методики и основные эксперименты по изучению мозга и поведения / Буреш Я., Бурешова О., Хьюстон Дж. П. – М.: Высш. шк., 1991. – 398 с.
204. Гаевый, М.Д. Использование гравитационных перегрузок в качестве скрининговой методики исследования новых биологически активных веществ / М.Д. Гаевый // Фармакология и токсикология. - 1986. - №3. – С. 101 – 102.
© 2010 Интернет База Рефератов