
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Дипломная работа: Особенности лихенофлоры сошненского лесничества Пинского района Брестской области
Дипломная работа: Особенности лихенофлоры сошненского лесничества Пинского района Брестской области
Министерство образования Республики Беларусь
УЧРЕЖДЕНИЕ ОБРАЗОВАНИЯ
«БРЕСТСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
ИМЕНИ А.С. ПУШКИНА»
БИОЛОГИЧЕСКИЙ ФАКУЛЬТЕТ
Кафедра ботаники и экологии
Дипломная работа
ОСОБЕННОСТИ ЛИХЕНОФЛОРЫ СОШНЕНСКОГО ЛЕСНИЧЕСТВА ПИНСКОГО РАЙОНА БРЕСТСКОЙ ОБЛАСТИ
Данилович Ольга Владимировна
студентка 5 курса специальности «Биология»
Научный руководитель:
Рецензент:
Допущено к защите
«__»____________2010 г.
Зав. Кафедрой ботаники и экологии
_________________
Брест 2010
СОДЕРЖАНИЕ
ВВЕДЕНИЕ
1 ОБЗОР ЛИТЕРАТУРЫ
1.1 Этимология названия
1.2 История исследования, систематическое положение
1.3 Микобионт, фотобионт и их симбиоз
1.4 Строение лишайников
1.4.1 Внешнее строение таллома
1.4.2 Грибы и водоросли, входящие в состав таллома лишайников
1.4.3 Анатомическое строение таллома
1.4.4 Органы спороношения лишайникового гриба
1.5 Размножение лишайников
1.6 Распространение и экологические группы лишайников
1.7 Хозяйственной значение лишайников
2 ОБЪЕКТ И МЕТОДЫ ИССЛЕДОВАНИЯ
2.1 Географическое положение и природные условия региона
2.2 Методика сбора полевого материала и последующая его обработка
3 РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
3.1 Растительность Пинского лесхоза
3.2 Систематическая характеристика собранных видов
ЗАКЛЮЧЕНИЕ
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
ВВЕДЕНИЕ
Проблема рационального использования природных ресурсов и их охраны может быть решена лишь комплексно совместными усилиями ученых и практиков на основе изучения и учета взаимосвязей и обусловленности многообразных природных процессов. Значительный интерес в этом плане представляют исследования в такой своеобразной группе растений, как лишайники: их видового состава, распространения, роли в различных биогеоценозах [1].
Лишайники относятся к сравнительно мало исследованным растительным организмам. Они представляют собой весьма своеобразную группу споровых растений, состоящую из двух компонентов гриба и одноклеточной, реже нитчатой водоросли, живущих совместно и образующих один целостный организм, обладающий иными, чем гриб и водоросль отдельно, морфологическими, анатомическими, физиологическими и экологическими свойствами. Известно более 20 000 видов и свыше 400 родов лишайников, встречающихся почти во всех природных зонах земного шара. Изучение их представляет теоретический и практический интерес.
Лишайники источник различных ценных медицинских препаратов и продуктов промышленного производства. Однако в настоящее время, в связи с недостаточностью их изученности, лишайники еще слабо используются.
Лишайники чутко реагируют на характер и состав субстрата, являясь хорошими индикаторами почвенно-грунтовых и микроклиматических условий, степени загрязненности воздуха в городах и населенных пунктах.
Лишайники могут быть использованы для датировки возраста различных предметов на основе измерения их слоевищ в диапазоне от нескольких десятилетий до нескольких тысячелетий [2].
Лишайники широко распространены в различных наземных биогеоценозах, но роль их в условиях Белоруссии особенно заметна в лесных сообществах. Здесь лишайники образуют группировки с большей биомассой, существенно влияют на жизнь леса [1].
Видовой состав лишайников очень разнообразен. Необходимость изучения лишайниковой флоры очевидна. В связи с этим, цель нашей работы на данном этапе – изучить видовой состав лишайников различных экологических групп территории Сошненского лесничества, одного из фоновых лесничеств Пинского лесхоза, где подобные исследования вообще не проводились.
Задачи исследования:
1. Собрать лишайники с различных субстратов в разных фитоценозах;
2. Определить до вида и их систематизировать;
3. Проанализировать информацию о собранных лишайниках;
4. Составить гербарий наиболее часто встречаемых видов.
5. Полученную информацию передать в лесхоз.
1. ОБЗОР ЛИТЕРАТУРЫ
Лиша́йники (лат. Lichenes) симбиотические ассоциации грибов (микобионт) и микроскопических зелёных водорослей и/или цианобактерий (фотобионт); микобионт образует слоевище (таллом), внутри которого располагаются клетки фотобионта.
1.1 Этимология названия
Русское название лишайники получили за визуальное сходство с проявлениями некоторых кожных заболеваний, получивших общее название «лишаи». Латинское название происходит от греческого λειχην (лат. Lichen) и переводится как бородавка, что связано с характерной формой плодовых тел некоторых представителей.
1.2 История исследования, систематическое положение
Usnea один из двух родов лишайников, описанных Теофрастом более двух тысяч лет назад. Первые описания известны из «Истории растений» Теофраста, который указал два лишайника – Usnea и Rocella, которую уже тогда использовали для получения красящих веществ. Теофраст предполагал, что они представляют собой наросты деревьев или водоросли. В XVII веке было известно только 28 видов.
Французский врач и ботаник Жозеф Питтон де Турнефор (рис. 1) в своей системе выделил лишайники в отдельную группу в составе мхов. Хотя к 1753 году было известно свыше 170 видов, Карл Линней описал только 80, охарактеризовав их как «скудное крестьянство растительности», и включил вместе с печёночниками в состав «наземных водорослей».
Началом лихенологии (науки о лишайниках) принято считать 1803 год, когда ученик Карла Линнея Эрик Ахариус (рис. 2) опубликовал свой труд «Methodus, qua omnes detectos lichenes ad genera redigere tentavit» («Методы, с помощью которых каждый сможет определять лишайники»). Он выделил их в самостоятельную группу и создал систему, основанную на строении плодовых тел, в которую вошли 906 описанных на то время видов.
Первым на симбиотическую природу в 1866 году на примере одного из видов указал врач и миколог Антон де Бари. В 1869 году ботаник Симон Швенденер (рис. 3) распространил эти представления на все виды. В том же году русские ботаники Андрей Сергеевич Фаминцин и Осип Васильевич Баранецкий обнаружили, что зеленые клетки в лишайнике – одноклеточные водоросли. Эти открытия были восприняты современниками как «удивительнейшие».
Сегодня лихенология является самостоятельной дисциплиной, смежной с микологией и ботаникой.
Традиционная систематика лишайников, оказывается во многом условна и отражает скорее особенности их строения и экологии, чем родственные отношения внутри группы, тем более что основывается она только на микобионте, а фотобионт сохраняет свою таксономическую самостоятельность.
Классифицируют лишайники по-разному, но в настоящее время рассматривают их как экологическую группу, уже не придавая им статуса таксона, поскольку независимость происхождения разных групп лишайников не вызывает сомнений, а группы, входящие в состав лишайников, помещают туда же, что и родственные микобионту грибы, не образующие лишайников. Для обозначения лишайников используют биноминальную номенклатуру, названия соотвествуют названию микобионта [3].

Рисунок 1 – Жозеф Питтон де Турнефор – впервые объединил лишайники в отдельную группу (в составе мхов).

Рисунок 2 – Эрик Ахариус – «отец лихенологии», выделил лишайники в самостоятельную группу и впервые систематизировал их.

Рисунок 3 – Симон Швенденер – первооткрыватель симбиотической природы лишайников.
1.3 Микобионт, фотобионт и их симбиоз
Лишайники очень интересная и своеобразная группа низших растений.
В лишайнике сочетаются два организма с противоположными свойствами: водоросль (чаще зеленая), которая в процессе фотосинтеза создает органическое вещество, и гриб, потребляющий это вещество. Взаимоотношения их строятся следующим образом. Гриб получает от водоросли органические вещества – углеводы, но в то же время как бы предоставляет водоросли, находящейся внутри тела лишайника, среду обитания, защиту от пересыхания и перегревания и т. д. Гриб снабжает водоросль достаточным количеством воды и растворенных в ней минеральных солей, которые он сам поглощает из окружающей среды (субстрата, атмосферного воздуха). Таким образом, хотя гриб в некоторой степени паразитирует на водоросли, но и она извлекает из совместной жизни с ним определенную пользу. Следовательно, в этом сожительстве наряду с паразитизмом имеются и черты симбиоза. В результате этих сложных взаимоотношений в процессе эволюции возник новый самостоятельный комплексный организм, имеющий новые, только ему присущие особенности строения и образа жизни и свои закономерности развития. При этом водоросль, выделенная из лишайника, в благоприятных условиях чаще всего может и вне его расти и развиваться самостоятельно, гриб же в этом случае обычно быстро погибает, так как в процессе приспособления к совместному существованию почти полностью потерял способность к самостоятельному развитию.
Поскольку основным формообразующим компонентом лишайников является гриб, то в последнее десятилетие среди ботаников получила распространение точка зрения, рассматривающая лишайники не как самостоятельную группу растений, а как грибы, включающие в свое тело водоросли, – так называемые лихенизированные грибы, которые перешли к паразитизму на водорослях и в связи с этим значительно изменились. При таком подходе виды лишайников распределяются среди грибов в соответствии с типом плодового тела лишайникового гриба и особенностями его строения. Однако своеобразная форма тела лишайников, присутствие особых веществ, неизвестных ни у грибов, ни у водорослей, особенности обмена веществ и ряд других признаков позволяют рассматривать лишайники как самостоятельную группу организмов [4].
1.4 Строение лишайников
1.4.1 Внешнее строение таллома
По форме и величине лишайники разнообразны, их размеры – от нескольких до десятков сантиметров. Вегетативное тело лишайников представлено слоевищем и в зависимости от образующегося в них пигмента может быть серым, сизым, зеленоватым, буро-коричневым, желтым, оранжевым или почти черным [5]. Пигменты лишайников откладываются в оболочках грибных гиф, реже в протоплазме. На интенсивность окраски сильное влияние оказывает освещенность: чем больше освещенность, тем ярче окраска.
Наиболее яркая окраска у лишайников высокогорных и приполярных районов Арктики и Антарктики. Это связано с тем, что пигменты, словно светофильтры, защищают лишайники от жесткого ультрафиолета, а в приполярных районах темноокрашенные лишайники быстрее нагреваются, снег и иней на их поверхности тает и превращается в капельножидкую влагу, необходимую лишайникам [6].
По внешнему виду различают три типа талломов лишайников: накипные (или корковые), листоватые и кустистые; эти типы связаны между собой переходными формами. Кроме них нередко различают еще чешуйчатый и филаментозный (нитевидный) типы талломов [7].
Таллом накипных лишайников представляет собой корочку, очень прочно срастающуюся с субстратом – корой деревьев, обнаженной древесиной, поверхностью скал и камней. Этот таллом невозможно отделить от субстрата, на котором он растет, не повредив его. Поверхность такого накипного таллома может быть порошковатой, зернистой, бугорчатой или реже гладкой; окраска – различной, обычно неяркой. У некоторых накипных лишайников почти весь таллом врастает в субстрат и на поверхности видны только плодовые тела лишайникового гриба; чаще это блюдцевидные апотеции.
Листоватые лишайники имеют вид чешуек или довольно крупных пластинок. Их таллом прикрепляется к субстрату обычно на большей своей части с помощью пучков грибных нитей (гиф) – ризин или отдельных тонких гиф – ризоидов. Лишь у немногих листоватых лишайников таллом срастается с субстратом только в одном месте с помощью мощного пучка грибных гиф, называемого гомфом. Между накипными и листоватыми лишайниками есть промежуточные формы, у которых таллом в центре накипной, а по краю листоватый, отстающий от субстрата.
У кустистых лишайников таллом состоит из ветвей или более толстых, часто ветвящихся стволиков. Такой кустистый лишайник срастается с субстратом только своим гомфом и растет вертикально либо наискось вверх (напочвенные виды) или свисая вниз (виды, растущие на стволах и ветвях деревьев). Между кустистыми и листоватыми талломами могут быть и переходные формы.
Для некоторых кустистых лишайников характерен так называемый первичный таллом, накипный или чаще листоватый, состоящий из мелких чешуек. На первичном талломе уже развивается вторичный, собственно кустистый, таллом в виде отдельных неразветвленных или разветвленных, вертикально стоящих веточек (рис. 4). Эти ветви вторичного таллома называются подециями и особенно характерны для рода кладония (С1аdоniа Hill. em. Vain.), у которого они очень разнообразны по форме. Подеции могут быть палочковидные, шиловидные, туповатые вверху или расширенные в виде бокальчика (сцифовидные), в виде кустиков с обильно разветвленными ветвями (рис. 4, 2). Сцифовидиые подеции часто пролиферируют, т. е. из центра сцифы или по ее краям вырастают новые сцифовидные подеции. У лишайников из родов кладония и стереокаулон (Stеrеосaulon Schreb.) на подециях развиваются чешуйки – филлокладии. Они соответствуют по происхождению чешуйкам первичного таллома, как бы переходящим на подеции (рис. 4, 3). Первичный таллом может сохраняться в течение всей жизни лишайника, но у ряда видов он быстро исчезает и остаются только подеции.
Талломы большинства кустистых лишайников имеют радиальное строение (у кладонии, стереокаулона, уснеи, алектории). Есть кустистые лишайники с лентовидным ветвящимся талломом, где морфологически хорошо различаются две стороны верхняя и нижняя (так называемое дорзовентральное строение). Такой таллом характерен для рода цетрария (Сеtraria Ach.) [4].

Рисунок 4 – Таллом кладоний: 1 – таллом (а – первичный, б – вторичный, или подеций), 2 – подеции кладоний (а, б – шиловидные, или палочковидные, в – с апотециями, г – сцифовидный с пролиферацией по краю, е – сцифовидный с пролиферацией в центре, ж кустистый), 3 – талломы с филлокладиями, 4 – филлокладии [а дланевидно(пальчато) разветвленные, б – бородавчатые (бугорчатые), в коралловидно разветвленные, г – щитовидно-чешуйчатые].
1.4.2 Грибы и водоросли, входящие в состав таллома лишайников
Среди лихенизированных грибов 90% связаны с зелеными водорослями, способными фиксировать атмосферный азот, а остальные 10% - с сине-зелеными водорослями из родов Nostoc Adanson, Scytonema Ag., Stigonema Ag., Ehichothrix Ag. и Calothrix Ag., усваивающими азот из атмосферы. Удивительно, что эти лишайники не характерны для местообитаний с недостатком азота, хотя водоросли в составе их таллома, безусловно, его фиксируют и высвобождают в значительном количестве [8].
Из одноклеточных зеленых водорослей в лишайниках встречаются виды родов хлорелла (Сhlоrеllа Beyer), цистококк (Сystoсоссиs Nüg); из нитчатых зеленых водорослей – кладофора (С1аdорhоrа Kütz), плеврококк (Рleuroсоссus auct.), трептеполия (Тrеntероhliа Mart.). При этом нити плеврококка и трентеполии в талломе лишайника часто распадаются на отдельные клетки. Виды этих родов – обычные, широко распространенные водоросли, часто встречающиеся как свободноживущие в пресных водоемах или па стволах деревьев. Нитчатая зеленая водоросль требуксия (Тrеbоихiа Puym.) живет только в талломах лишайников. Из сине-зеленых водорослей в талломах лишайников чаще всего встречаются представители рода носток (Nostoс Adanson); нитчатое тело этих водорослей в талломе лишайника также обычно распадается на отдельные участки. Водоросль, входящая в состав таллома лишайника, называется фотобионт.
Грибы, входящие в состав лишайников, в основном относятся к классу сумчатых (Аsсотусеtеs). Гифы гриба в подавляющем большинстве случаев (за исключением слизистых лишайников) составляют основу таллома лишайников, водоросль же как бы заключена между гифами гриба. Гриб, входящий в состав таллома лишайника, называется микобионт [4]
1.4.3 Анатомическое строение таллома
По анатомическому строению различают два типа талломов лишайников. В более примитивном, гомеомерном талломе клетки или нити водоросли более или менее равномерно распределены между гифами гриба по всей толщине таллома. Такой таллом имеют слизистые лишайники, содержащие в качестве фикобионта нитчатые сине-зеленые водоросли. Нити этих водорослей окружены большим количеством слизи, в которой по всем направлениям проходят грибные гифы, что хорошо видно на поперечном срезе таллома под микроскопом или через 10-кратную лупу. В сухом состоянии такие талломы имеют вид буроватых или черноватых морщинистых корочек. Во влажную погоду они быстро впитывают влагу, разбухают и приобретают форму листоватых лишайников [9]. Один из таких лишайников – коллема (Соllета Web.), виды которого широко распространены на скалах и камнях на Черноморском побережье Крыма, Кавказа и сопредельных областей. Это черноватые или темно-оливковые подушечки, образуемые извилистыми приподнятыми лопастями.
Более сложно построен гетеромерный таллом. На поперечном срезе такого таллома под микроскопом хорошо различимы элементы его строения. Верхняя кора таллома образована плотным переплетением гиф гриба. Дальше идет гонидиальный слой, состоящий из клеток водоросли. Гифы гриба, заходящие в гонидиальный слой, образуют мелкие разветвления, которые плотно примыкают к клеткам водоросли. Здесь гриб получает от фотосинтезирующей водоросли углеводы. Дальше расположена сердцевина (слой из рыхлопереплетенпых гиф гриба), с помощью, которой внутри таллома поддерживаются определенная влажность и воздушная среда, необходимая и для самих гиф, и для клеток водоросли. За сердцевиной расположена нижняя кора из плотно переплетенных гиф гриба. Гетеромерный таллом хорошо выражен у листоватых лишайников и у кустистых лишайников с лентовидным дорзовентральным талломом.
У кустистых лишайников с цилиндрическим радиальным талломом (сцифовидный, палочковидный и т. д.) таллом имеет гетеромерно-радиальное строение. Под корой, одевающей снаружи ветви такого таллома, лежит гонидиальный слой, идущий вокруг всего таллома, а внутрь от него расположена сердцевина. У накипных лишайников с гетеромерным талломом никогда не бывает нижней коры, они срастаются с субстратом сердцевинными гифами.
Развитие и выраженность слоев гетеромерного таллома у разных видов лишайников различны. У ряда листоватых и кустистых лишайников с гетеромерным талломом происходит местный разрыв корового слоя и возникают плоские, неотчетливо очерченные пятна, обычно более светлые, чем сама кора (макулы, или цифеллы); они служат для проведения воздуха в сердцевину таллома и у ряда видов являются определительным систематическим признаком. Встречаются макулы у рода цетрария, например у «исландского мха» (Сеtraria islandicа L.)
1.4.4 Органы спороношения лишайникового гриба
На талломе лишайника из грибных гиф формируются плодовые тела гриба со спорами. Это в основном расположенные на поверхности таллома апотеций или погруженные в таллом кувшиновидные перитеции.
В апотециях и перитециях формируются споры для размножения лишайникового гриба.
Перитеции встречаются у небольшого числа лишайников. Апотеции образуются у многих видов лишайников. Они имеют чаще блюдцевидную форму, но встречаются и более или менее выпуклые или почти шаровидные. Самые крупные апотеции имеют диаметр более 1 см, но лишайников с такими апотециями немного. У громадного большинства лишайников диаметр апотециев от одного до нескольких миллиметров. Они разбросаны по поверхности листоватого таллома, чаще в его середине, или расположены по краям его лопастей. У кустистых лишайников апотеции обычно находятся на концах веточек или сциф. Они бывают сидячими или реже на небольших ножках приподнимаются над талломом. Апотеции могут быть окрашены в один цвет с талломом, или поверхность апотеция (так называемый диск апотеция) имеет другую окраску.
По строению различают три типа апотециев: леканориновый, лецидеиновый и биаториновый.
Леканориновый апотеций по своему анатомическому строению сходен с талломом лишайника. Его диск имеет талломный (слоевищный) край, образованный талломом лишайника и состоящий из гиф гриба и клеток водоросли. Этот край окрашен так же, как и таллом лишайника, и отличается по окраске от самого диска. В самом талломе под леканориновый апотецием также имеются клетки водоросли (рис. 5, 2).
Лецидеиновый апотеций имеет край, состоящий только из гиф гриба и окрашенный в тот же цвет, что и диск апотеция. В этом крае, а также в талломе под лецидеиновым апотецием клеток водорослей нет. Лецидеиновые апотеции имеют твердую консистенцию и обычно темную окраску (рис. 5, 3).
Биаториновый апотеций имеет такое же строение, как лецидеиновый, но отличается яркой окраской и мягкой консистенцией.
1.5 Размножение лишайников
Споры лишайникового гриба, выбрасываемые из перитециев и апотециев, прорастают в гифы, которые, переплетаясь, образуют зачаточный таллом лишайника, или прототаллом. Чтобы в дальнейшем из него возник настоящий лишайник, необходим контакт прототаллома с определенным видом водоросли, а также определенные внешние условия. В противном случае прототаллом быстро погибает. Таким образом, споры гриба, образующиеся в апотециях и перитециях, большого значения для размножения лишайников не имеют, поскольку сочетание необходимых для этого условий в природе встречается редко.
Более важны для размножения лишайников такие образования, в которых одновременно присутствуют гифы гриба и клетки водоросли. Это соредии и изидии. Они служат для размножения лишайника как целого организма. Попав в благоприятные условия, они дают начало непосредственно новому таллому. Соредии и изидии встречаются чаще у листоватых и кустистых лишайников.
Соредии представляют собой мельчайшие образования в виде пылинок, состоящих из одной или нескольких клеток водоросли, окруженных гифами гриба (рис. 5, 4). Формирование их обычно начинается в гонидиальном слое. Вследствие массового образования соредии количество их увеличивается, они давят на верхнюю кору, разрывают ее и оказываются на поверхности таллома, откуда легко сдуваются при любом движении воздуха или смываются водой. Скопления соредии называют соралями. Наличие и отсутствие соредии и соралей, их расположение, форма и окраска постоянны для определенных лишайников и служат определительным признаком.
Иногда при отмирании лишайников их таллом превращается в порошковатую массу, состоящую из соредии. Это так называемые лепрозные формы лишайников (от греческого слова «лепрос» — «шероховатый», «неровный»). В этом случае определить лишайник почти не представляется возможным.
Соредии, разносимые ветром и дождевой водой, попав в благоприятные условия, постепенно образуют новый таллом. Возобновление нового таллома из соредии происходит очень медленно. Так, у видов из рода кладония нормальные чешуйки первичного таллома развиваются из соредии только через срок от 9 до 24 месяцев. А для развития вторичного таллома с апотециями требуется от одного до восьми лет в зависимости от вида лишайника и внешних условий.
Если условия для развития нового таллома не совсем благоприятны, но все же возможен рост грибных гиф и размножение водорослей, то из соредии развиваются новые соредии и возникает описанная выше лепрозная форма, имеющая вид порошковатого пятна.
Изидии встречаются у меньшего числа видов лишайников, нежели соредии и сорали. Они представляют собой простые или коралловидно разветвленные выросты, обычно густо покрывающие верхнюю сторону таллома. В отличие от соралей изидии снаружи покрыты корой, часто более темной, чем таллом. Внутри, под корой, они содержат водоросли и грибные гифы (рис. 5, 5). Изидии легко отламываются от поверхности таллома. Обламываясь и распространяясь с помощью дождя и ветра, они, так же как и соредии, могут при благоприятных условиях образовывать новые талломы лишайников.
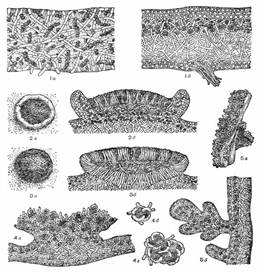
Рисунок 5 – 1 разрез таллома (а – гомеомерного, б – гетеромерного) 2 леканориновый апотеций (а – вид сверху, б – разрез), 3 лецидеиновый апотеций (а – вид сверху, б – разрез), 4 строение соралей (а – сораль, б, в – отдельные соредии), 5 изидии (а – внешний вид, б – разрез).
Многие лишайники не образуют апотециев, соредий и изидии и размножаются участками таллома, которые легко отламываются от хрупких в сухую погоду лишайников ветром или животными и ими же переносятся. Особенно широко распространено размножение лишайников участками таллома в арктических областях. В наибольшей степени размножение кусочками таллома присуще накипным эпилитным лишайникам. Появление трещин в скалах и камнях, покрытых лишайниками, а также появление трещин в самом талломе накипного лишайника из-за неодинакового отношения к нагреванию и увлажнению камня и самого лишайника ведет к разламыванию таллома и отделению небольших его участков. Эти отделившиеся кусочки таллома и переносятся ветром и водой на новые места, давая начало новым талломам.
Для некоторых лишайников (около 100 видов из разных родов) характерно наличие на поверхности, а иногда и внутри таллома образований разного размера в виде разбросанных небольших вздутий, бородавочек, булавовидных, шаровидных и коралловидных выростов. Это цефалодии. Они могут покрывать верхнюю или нижнюю сторону листоватых талломов или развиваться по бокам подециев либо веточек кустистых талломов. В отличие от изидии они более разнообразной формы и разбросаны более редко. На срезе под микроскопом видно, что они имеют гомеомерное строение и состоят из беспорядочного скопления гиф, среди которых равномерно распределены клетки или нити сине-зеленых водорослей. Реже цефалодии имеет гетеромерное строение. Водоросли в цефалодиях всегда сине-зеленые, тогда как в талломе лишайника, несущего цефалодии, водоросли всегда зеленые. Образование цефалодиев обусловлено проникновением в таллом не свойственных данному виду лишайника сине-зеленых водорослей, которые вызывают местное разрастание гиф гриба, «обрастающего» эти водоросли. Роль цефалодиев для самого лишайника не совсем ясна. Предполагается и отчасти доказано экспериментально, что они увеличивают питание лишайника, живущего на бедном субстрате. Сине-зеленые водоросли цефалодия способны фиксировать атмосферный азот, который затем в виде азотистых соединений усваивается всем талломом лишайника – составляющими его грибом и зеленой водорослью, которые сами не могут фиксировать атмосферный азот.
Цефалодии специфичны для ряда видов и имеют значение определительного (систематического) признака. Характерный пример лишайника с цефалодиями – напочвенный вид пельтигера пупырчатая (Peltigera aphtnosa (L.) Willd.).
1.6 Распространение и экологические группы лишайников
Лишайники насчитывают около 20 тысяч видов и широко распространены по земному шару – от полярных холодных скал до раскаленных камней пустынь.
Закономерности их географического распространения изучены еще недостаточно. С одной стороны, отмечается приуроченность определенных видов к тем или иным природным зонам, (например, есть виды, приуроченные к зоне таежных лесов, такие, как уснея длиннейшая – Usnea longissima Ach.). Но есть виды, которые в своем распространении связаны не столько с природными условиями определенной зоны, сколько с условиями, которые повторяются в нескольких природных зонах [виды, связанные с океаническим климатом, многие арктоальпийские виды и т. д.; стереокаулон альпийский (Stereocaulon alpinum Laur.), например, растет на Крайнем Севере на каменистой почве тундр, а также в альпийском поясе различных горных массивов (Алтай, Кавказ)]. Географическое распространение многих лишайников прямо связано с их выборочным отношением к субстрату, хотя последний и не является основной причиной, ограничивающей их распространение. Безразличные к субстратам виды лишайников имеют более широкий ареал (область распространения), чем виды, более требовательные к определенному субстрату. Есть лишайники космополиты, распространенные по всему земному шару, но таких относительно немного.
По приуроченности к субстрату лишайники подразделяют на несколько экологических групп.
Напочвенные, или эпигейные, лишайники. Виды этой группы должны выдерживать сильную конкуренцию со стороны быстрорастущих высших растений, особенно травянистых. Поэтому они редко встречаются на плодородных почвах и достигают большего развития в местах, мало пригодных для высших растений из-за незначительной питательности субстрата или неблагоприятных климатических условий, например на песчаных почвах, в тундрах, полупустынях, на торфяниках и т. д.
Напочвенные лишайники могут расти как на открытых местах, так и в лесах. Лишайники открытых пространств произрастают вдоль дорог, на старых пожарищах, на бедных сухих и болотистых лугах, на опушках. Это пельтигера собачья (Peltigera canina (L.) Willd.), пельтигера бородавчатая (P. aphthosa (L.) Willd.), пельтигера прорывающаяся (P. erumpens (L.) Willd.), «исландский мох» (Cetraria islandica (L.) Ach.), стереокаулон войлочный (Stereocaulon tomentosum Fr.), различные виды рода кладония (Cladonia Hill. em. Vain). Особенно пышно разрастаются напочвенные лишайники открытых пространств в тундре и лесотундре, где часто занимают громадные площади и определяют характер ландшафта. Это широко распространенные виды «оленьего мха»: кладония оленья (C. rangiferina (L.)), кладония приальпийская (C. alpestris (L.) Rabh.), кладония лесная (C. sylvatica f. tenius). Кроме них здесь растут различные кустистые виды рода цетрария (Cetraria Ach.), виды родов алектория (Alectoria Ach.), стереокаулон и др. Очень характерна для тундры охролехия двуполая (Ochrolechia androgyna (Hoffm.) Arn.), иногда на больших площадях образующая, беловатую корочку на мхах, некоторых цветковых растениях и даже на других лишайниках. Достигая большого развития, охролехия препятствует фотосинтезу и как бы удушает растения, на которых поселилась. Широко распространены в тундре также охролехия виннокаменная (O. tartarea (L.) Massal.) и охролехия холодная (O. frigid (Sw.) Lynge).
Напочвенные лишайники открытых пространств встречаются также в сухих степях и полупустынях, на скалах и каменных россыпях в высокогорных районах. Они образуют две четкие подгруппы видов. Первая подгруппа – «кочующие» лишайники, не срастающиеся с почвой и переносимые ветром с места на место, как перекати-поле. Это в основном мелколистоватые лишайники: аспицилия съедобная, или «лишайниковая манна» (Aspicilia esculenta (Pall.) Flag.), пармелия блуждающая (Parmelia vagans Nyl.), пармелия грубоморщинистая (P. ryssolea (Ach.) Nyl.), корникулярия степная (C. steppae Ach.). Вторая подгруппа – прикрепленные к почве лишайники; в основном это накипные виды, образующие в сухую погоду на почве малозаметные налеты или корочки. Среди них есть виды, приуроченные к песчаным [рамалина многообразная (Ramalina polymorpha (L.) Ach.), рамалина скрученная (R. strepsilis (L.) Ach.), пармелия темно-бурая (Parmelia pulla (L.) Ach.) и др.], известковым [виды родов веррукария (Verrucaria Wigg. em. Th. Fr.) и коллема (Collema Web.)] и глинистым [беомицес розовый (Baeomyces roseus Pers.) и беомицес рыжий (Baeomyces rufus (Huds.))] почвам.
Напочвенные лишайники в лесу, особенно негустом, представлены различными видами кладоний, пельтигер. Особенно большого развития лишайниковый покров достигает в сухих сосняках. Это так называемые боры-беломошники. Покров из кладоний бывает сильно развит и в лиственничных лесах Восточной Сибири.
К представителям напочвенных лишайников лесов близко примыкают по своей экологии лишайники, развивающиеся на гниющих пнях и на основаниях стволов деревьев. В большинстве своем они теневыносливы и влаголюбивы. Это цетрария сосновая (Cetraria pinastri (Scop.) Röhl.), кладония пальчатая (Cladonia digitata Schaer.), упомянутые уже виды кладоний, составляющие «олений мох», и т. д. Эти виды являются переходными к следующей экологической группе.
Эпифитные лишайники поселяются на деревьях и кустарниках. Среди них также можно выделить несколько подгрупп: эпифилльные лишайники, растущие на листьях деревьев и кустарников; настоящие эпифитные лишайники, растущие на коре; эпиксильные лишайники, растущие на обнаженной и обработанной древесине.
Эпифилльные лишайники сравнительно немногочисленны. Распространены они главным образом в тропиках и отчасти в субтропиках на старых листьях вечнозеленых растений. Большей частью они используют зеленый лист только как опорный субстрат. Эпифилльные лишайники живут в основном на поверхности листа, не проникая внутрь него и не причиняя ему вреда. Но есть виды, которые проникают под кутикулу и прикрепляются к наружным стенкам клеток эпидермиса листа. Эпидермис от этого или страдает мало, или отмирает, и в этом случае на листьях обычно образуются серые или серо-зеленые пятна. На территории бывшего СССР немногочисленные эпифилльные лишайники встречаются в основном на юге Черноморского побережья Кавказа. Из них можно указать на катилларию Боутелли (Catilaria bouteillei Mass. em. Th. Fr.) и бацидию грушевидную (Bacidia apiahica Koerb.), растущие на верхней поверхности старых листьев чайного куста, что, без сомнения, отражается на общем развитии куста. Катиллария Боутелли встречается также на хвое и живых веточках елей и пихт.
Эпифилльными, хотя и с известной приблизительностью, можно назвать некоторые распространенные листоватые лишайники из родов фисция (Physcia (Schreb.) Vain) и пармелия (Раrmelia Ach.), иногда нарастающие с коры веток на хвою хвойных деревьев.
Эпифитные лишайники на коре деревьев очень многочисленны. Здесь обитают и накипные, и листоватые, и кустистые формы. Нередко они сплошь покрывают ствол дерева на большом протяжении. На участке коры величиной не более ладони иногда насчитывали до 38 видов лишайников, которые росли вплотную друг около друга и даже один на другом.
Существует ли какая-нибудь зависимость флоры лишайников от древесной породы, еще точно не установлено. Правда, наблюдения показывают, что на отдельных породах деревьев часто наблюдаются определенные группировки лишайников. Так, в Подмосковье на осинах чаще встречаются стенная золотнянка (Xanthoria parietina Belt.), фисция припудренная (Physcia pulverulenta (Schreb.) Hampe.), фисция щетинистая (Ph. hispida (Schreb.) Fr.), фисция звездчатая (Рh. stellaris (Ach.) Nyl.), фисция темная, или реснитчатая (Рh. ciliate (Hoffm.) DR.), пармелия бороздчатая (Раrmelia sulcata Tayl.); на сосне – уснея длиннейшая (Usnea longissima Ach.), алектория оливковая (Alectoria olivacea Ach.), рамалина волосовидная (Ramalina thrausta (Ach.) Nyl.); на рябине и дубе – пармелия бороздчатая. Эти же виды могут встречаться и на других породах, хотя и не столь часто.
Для преобладающего расселения того или иного вида лишайников имеет значение строение коры (ее физические свойства, химический состав, кислотность и т. д.). Приуроченность лишайников к определенным древесным породам в известной степени зависит и от климатических условий, в которых произрастает данная порода. Например, различаются по качественному и количественному составу лишайников сосны Прибалтики, средней полосы Европейской части СССР и Европейского Севера.
Распространено мнение, что эпифитные лишайники поселяются на старых, ослабленных деревьях. Действительно, в ряде случаев это так: на старых экземплярах елей более богатый видовой состав лишайников; старые, ослабленные плохим уходом и нарушением агротехники кусты чая также быстрее заселяются эпифилльными лишайниками. Однако лишайники часто растут и на молодых, хорошо развитых деревьях и кустарниках [4].
Расселение лишайников на стволе зависит в основном от освещенности. Лишайники, приспособленные к существованию при малой освещенности, поселяются ближе к основанию ствола, а светолюбивые поднимаются по стволу. На основании ствола лишайники конкурируют с мхами. Вероятно, здесь они частично паразитируют на мхах [10].
Флора эпиксильных лишайников, поселяющихся на обнаженной или обработанной древесине, обычно близка по видовому составу к флоре лишайников, обитающих па коре деревьев, почве и отчасти на скалах.
Эпилитные лишайники поселяются; на камнях и скалах и представлены в основном накипными видами. Расселение их по субстрату различно. Один вид может сплошь покрывать скалу или крупный камень на большой площади, придавая им заметную издали, характерную для определенного вида лишайника желтую, оранжевую, зеленоватую, коричневатую, черную или другую окраску. В другом случае на небольшом участке могут произрастать несколько видов лишайников, образуя на субстрате благодаря своей разнообразной окраске пестрый узорчатый рисунок.
Среди эпилитных лишайников различаются кальцефильные виды, живущие на известковых породах [виды рода веррукария (Verrucaria Wigg. em. Th. Fr.), коллема черная (Collema multifidum (Huds.) DC.), гаcпарриния обманчивая (Gasparinia decipiens (Arn.) Sydow.), некоторые виды рода биатора (Вiotora Ach. em. Koerb.) и др.], и кальцефобные, живущие па кремнеземных породах [виды родов лецидея (Lecidea Ach.), леканора (Lecanora (Ach.) Th. Fr.), ризокарпон географический (Rhizocarpon geographycum (L.) DC.)]. Имеются также индифферентные виды, поселяющиеся и на тех, и на других породах. Среди лишайников, живущих на кремнеземных породах, некоторые селятся преимущественно на кислых (порфир), другие – преимущественно на основных (базальт, диабаз) породах. На горных породах, содержащих различные минералы, например на граните, лишайники сначала заселяют более мягкую часть – слюду и уже позднее – кварц. При первоначальном заселении используются все выветрившиеся места и мельчайшие трещинки в горной породе, в которые легче могут проникнуть грибные гифы. Очевидно, к эпилитным нужно отнести и лишайники, поселяющиеся на кирпичных строениях, черепичных крышах и т. д.
Особую экологическую группу составляют очень своеобразные водные лишайники, постоянно или большую часть года проводящие под водой. Эти лишайники биологически мало изучены. Среди них есть виды пресноводные и обитающие в соленой воде, они могут быть обитателями стоячих водоемов и быстро текущих рек и ручьев. У водных лишайников есть некоторые приспособления к обитанию в воде, сводящиеся к защите плодового тела лишайникового гриба. Обычно оно бывает глубоко погружено в таллом лишайника и одето плотной оболочкой. Настоящие подводные лишайники обычно селятся в прозрачной, чистой воде и заходят на несколько метров вглубь. Дерматокарпон речной (Dermatocarpon aquaticum (Weiss.) Zahlbr.) поселяется на камнях и скалистом ложе ручьев, речек и светлых озер Карелии [11].
Между собственно водными и наземными видами есть ряд переходных по обитанию видов. Они то живут под водой, то селятся на периодически омываемых водой местах, то только близ воды, довольствуясь в этом случае испаряющейся влагой, брызгами от водопадов или прибоя. Наконец, есть виды, которые могут выдерживать длительное случайное затопление, но обычно живут вне воды. Это ризокарпон темный (Rhizocarpon obscuratum (L.) DC), лецидея беловато-голубоватая (Lecidea albocoerulescens (Ach.)) и др.
Названные основные четыре группы (напочвенные, эпифитные, эпилитные и водные) не исчерпывают экологического разнообразия лишайников. Лишайники могут встречаться в различных местах и на всевозможных субстратах, казалось бы совсем не подходящих для обитания. Их находили на железе (на старых пушках), на смоле, древесном угле, костях, оконных стеклах, фаянсе, фарфоре, коже, картоне, линолеуме, войлоке, различных материях и т. д. Это разнообразие субстратов показывает, что лишайники часто используют субстрат лишь как опору [4].
Лишайники очень медленно растущие организмы. Прирост их таллома при благоприятных условиях колеблется в зависимости от вида от 1 до 8 мм в год. При этом листоватые и кустистые лишайники растут быстрее, чем накипные. Средний возраст лишайников – от 30 до 80 лет, а отдельные экземпляры, как можно определить по косвенным данным, доживают до 600 лет. Имеются указания, что иные талломы лишайников насчитывают даже около 2000 лет. Таким образом, наряду с гигантской секвойей и остистой сосной лишайники можно считать самыми долгоживущими организмами. Нужно отметить, что ввиду медленного роста лишайников необходимое условие их жизни – продолжительная неподвижность субстрата.
В растительном покрове лишайники, как и мохообразные, играют значительную роль, особенно в северных и высокогорных областях. Они существенные компоненты растительного покрова равнинных и горных тундр и отчасти таежных лесов. Вытекающие из специфической двойной природы лишайников преимущества позволяют им расти в таких условиях, в которых не могут существовать другие организмы. Так, лишайники являются пионерами зарастания каменистых субстратов; осваивая их, они прокладывают дорогу другим растениям [12].
Хорошо известна способность лишайников переносить длительное промерзание зимой в состоянии анабиоза. В одном из опытов лишайник Cladonia Hill. em. Vain. замораживали при -15°С на 110 недель (более двух лет). После оттаивания лишайник оказался живым и вполне жизнеспособным, у него возобновились фотосинтез и рост. Очевидно, у лишайников в крайне холодных условиях существования периоды такого анабиоза очень длительны, а рост и активная жизнедеятельность осуществляется лишь в короткие благоприятные периоды (причем не каждый год). Такое частое прерывание активной жизни на долгие сроки, по-видимому, объясняет колоссальный возраст многих лишайников, определенный радиоуглеродным методом (до 1300 лет у Rhizocarpon geographicum (L.) DC. в Альпах, до 4500 лет у лишайников в Западной Гренландии) [13].
1.7 Хозяйственной значение лишайников
Будучи одним из основных напочвенных растений тундровой зоны, некоторые лишайники (кладонии и цетрарии) являются главным кормовым растением для северных оленей. Олени обладают способностью чувствовать запах лишайников и сквозь снеговой покров, из-под которого они их достают, разгребая снег копытами. При кормежке олени скусывают лишь верхушки кустиков кладоний и цетрарий, и лишайники продолжают свой рост. Однако растут они медленно, и для восстановления пастбищ требуется от 10 до 30 лет. Лишайники используют в пищу и другие животные; например, свиньи и овцы охотно поедают кладонию листоватую, кладонию оленевидную и др.
В Японии употребляется в пищу и даже служит предметом экспорта в страны Юго-Восточной Азии растущий на скалах листоватый лишайник гирофора съедобная (Gygrophora esculenta Ach.). Съедобен и лишайник аспицилия съедобная (Aspicilia esculenta (Pall.) Flag.), растущий в степной и полупустынной зонах. Он содержит до 55-65% щавелевокислого кальция. Однако усвояемость питательных веществ лишайников очень незначительна.
В результате взаимодействия гриба и водоросли, составляющих организм лишайника, образуются специфические вещества, которые в природе нигде больше не встречаются. Это так называемые лишайниковые вещества, или лишайниковые кислоты. Некоторые из этих кислот обладают антибиотическим действием, например усниновая кислота, образуемая 70 видами лишайников. Это сильный антибиотик, который под названием «бинан» введен в медицинскую практику для лечения некоторых заболеваний и рекомендован к применению в ветеринарии [4]. Основные органические вещества, накапливающихся в большинстве лишайников, – полисахариды (в частности, лихенин, но у некоторых вырабатывается полуклетчатка). Белков и жиров содержится мало. В лишайниках имеется много железа, аммония, кремнекислоты [14]. Некоторые лишайниковые вещества действуют как стимуляторы, поднимающие тонус организма. На этом основано использование в народной медицине отваров цетрарии исландской («исландский мох»). В ее состав входит паралихестериновая кислота, обладающая тонизирующим действием. Из широко распространенного лишайника эверния сливовая («дубовый мох») извлечено вещество резиноид обладающее ароматическими свойствами и являющееся хорошим закрепителем аромата. Этот препарат заменяет соответствующее импортное сырье для парфюмерной промышленности. К духам, в состав которых входит резиноид, относятся «Шипр», «Кристалл», «Кремль», «Кармен», «Маска», «Свежее сено» и др. Этот же лишайник и реже другой вид эверния шелушащаяся – применялись в странах Северной Африки для ароматизации хлеба. Некоторые лишайники из рода рочелла, растущие на морских побережьях, а также охролехия виннокаменная, растущая на скалах и на почве в северных районах СССР, применяются местным населением как красители [4].
Среди двадцати тысяч видов лишайников наибольшее практическое значение имеет ягель, который на Руси издавна использовали для выпечки хлеба в голодные годы. Крестьяне вымачивали ягель в течение суток в растворе соды для удаления горького вкуса, затем тщательно промывали водой, сушили и измельчали в муку.
Ещё в XIX столетии европейские врачи широко рекомендовали ягель для лечения туберкулёза и коклюша. Позднее увлечение ягелем прошло и лишь сравнительно недавно забытый лишайник вновь заинтересовал учёных, поскольку химикам удалось выделить из него усниновую кислоту — могучий антибиотик, убивающий наружболезнетворные микроорганизмы в ничтожно малых концентрациях. Препараты усниновой кислоты на спирту или в масляном растворе применяют наружно для лечения гнойных ран и ожогов.
Иногда ягель неправильно называют «исландским мхом» [15].
Система наблюдения за реакцией биологических объектов на воздействие загрязнителей называется биологическим мониторингом. Биологический мониторинг включает в себя наблюдение, оценку и прогноз изменения состояния экосистем и их элементов, вызываемого антропогенным воздействием. Идеальная система мониторинга дает возможность количественно оценить состояние среды и ее изменения. Биологическая индикация позволяет оценивать степень загрязнения окружающей среды по существующим биологическим показателям. Поскольку вариабельность органической жизни чрезвычайно велика, особенно важно выбрать такие организмы, которые являются лучшими индикаторами трендов изменения среды [16].
Одними из наиболее известных биологических индикаторов являются лишайники, чувствительность которых обусловлена их физиологией и симбиотической природой. Лишайники выбраны объектом глобального биологического мониторинга, поскольку они распространены по всему Земному шару и поскольку их реакция на внешнее воздействие очень сильна, а собственная изменчивость незначительна по сравнению с другими организмами. Из всех экологических групп лишайников наибольшей чувствительностью обладают лишайники-эпифиты [17].
В перспективе при широком и углубленном изучении лишайники могут стать источниками ценных биологически активных веществ (медицинских препаратов и т. д.). Однако использование лишайников должно вестись планомерно, на заранее разработанной научной основе, чтобы не нанести непоправимый ущерб природе, в которой они играют свою определенную важную роль [4].
2. ОБЪЕКТ И МЕТОДЫ ИССЛЕДОВАНИЯ
2.1 Географическое положение и природные условия региона
Лишайники, в отличие от высших растений, можно собирать для гербария круглый год. Сбор материала для дипломной работы производился на территории Сошненского лесничества Пинского лесхоза. Пинский лесхоз Брестского производственного лесохозяйственного объединения Министерства лесного хозяйства Республики Беларусь расположен в южной части Брестской области Республики Беларусь на территории двух административных районов: Пинского (52,4%) и Ивановского (47,6%). Протяженность территории лесхоза с севера на юг – 55 км, с запада на восток – 91 км.
Климат района расположения лесхоза теплый, неустойчиво-влажный атлантико-континентальной области. Среднегодовая температура воздуха по метеоданным многолетних наблюдений составляет 6,8°С, среднегодовое количество осадков 590 мм, причем их максимум приходится на теплый период года. Зима умеренно-холодная общей продолжительностью около 4 месяцев. Переход температуры через плюс 10° происходит весной в период с 20 по 25 апреля и осенью – с 10 по 15 октября. Среднегодовая продолжительность вегетационного периода – около 6,5 месяцев.
В целом же климат района можно охарактеризовать как умеренно теплый с достаточным количеством выпадающих осадков, продолжительным вегетационным периодом, относительно нехолодной зимой и теплым летом. Природно-климатические условия вполне благоприятны для произрастания основных лесообразующих пород. Вполне приемлемы климатические условия и для произрастания твердолиственных пород (дуба, клена, ясеня).
Согласно геоморфологическому районированию Беларуси территория лесхоза относится к Полесской низменности. Рельеф этой территории в целом равнинный, пониженный. Преобладающие высоты достигают 134-147 метров над уровнем моря.
В технологическом отношении основная часть территории лесхоза приурочена к Полесской седловине с глубиной залегания фундамента 60-180 м, изредка 200-300 м.
Четвертичные отложения имеют мощность до 60-70 м в западной части лесхоза и 30-40 м в юго-восточной части. Представлены, в основном, водно-ледниковыми и древнеаллювиальными отложениями эпохи днепровского оледенения на территории Загородья, междуречье Пины и Ясельды, древнеаллювиальными и болотными отложениями валдайского и голоценового возраста на остальной территории.
В 1990-91 годах I-й Минской лесоустроительной экспедицией ПО «Белгослес» было проведено почвенное обследование территории лесхоза. Исходя из результатов этого обследования следует, что по типу почвообразовательного процесса в лесхозе абсолютно преобладают дерново-подзолистые почвы – 73,8 %. На долю торфяно-болотных, пойменных и дерновых заболачиваемых почв приходится 26,2 % территории лесхоза. Автоморфные почвы (нормального увлажнения) занимают 7,9 % территории лесхоза. Остальная часть территории в различной степени переувлажнена: от полугидроморфных (близких к нормальной степени увлажнения) – 74,5 % до гидроморфных – 17,6 %. По механическому составу наибольшее распространение имеют бедные по плодородию песчаные почвы, удельный вес которых составляет 76,6 %. Супесчаные почвы занимают 4,1 %, а перегнойные – 1,7%. Заболоченные почвы занимают 26,2 % территории лесхоза.
Преобладающие на территории лесхоза полугидроморфные дерново-подзолистые песчаные почвы обладают очень высокой водопроницаемостью, вследствие чего карбонаты и полуторные окислы в них вмыты, как правило, на очень большую глубину. Почвы эти бедны элементами питания для растений. На ровных участках поверхности пески имеют связный механический состав и развивающиеся на них почвы являются более плодородными. Здесь обычно формируются сосновые древостои I-II бонитетов, часто с примесью березы и дуба. Дерново-подзолистые супесчаные почвы встречаются реже. Они значительно богаче элементами питания и заняты высокопродуктивными смешанными дубравами, сосняками и их производными. Дерново- и торфяно-болотные почвы на территории лесхоза распространены весьма широко. Развиваются они на понижениях рельефа в связи с избытком увлажнения. В долинах рек и ручьев, где широко выражено проточное увлажнение, произрастают высокопроизводительные черноольховые насаждения.
Все почвы лесхоза по степени кислотности разделены на 6 групп – от очень сильнокислых до нейтральных. Абсолютно преобладают почвы сильнокислые и очень сильнокислые.
По гидрологическому районированию территория лесхоза относится к Днепровскому гидрологичнскому району. Основным водоприемником для всех речек и ручьев, протекающих на его территории, является р. Припять – крупнейший приток Днепра.
2.2 Методика сбора полевого материала и последующая его обработка
Сбор полевого материала производился методом рекогнасцировки местности. Пользуясь определителем, идентифицировались собранные нами виды. Для работы наиболее удобны «Лишайники Белоруссии. Определитель» (Горбач Н. В., 1973), «Водоросли, лишайники и мохообразные СССР» (Горленко М. В., 1978), «Лишайники, бактерии и актиномицеты» (Комарницкий, 1960). Определение лишайников проводилось с помощью реактива 10%-го водного раствора КОН.
Собранные лишайники упаковывали сразу на месте. Для этого их вкладывали в заранее заготовленные пакеты (конверты) таким образом, чтобы в один пакет помещались лишайники только из одного местообитания – с одного участка ствола, скалы и т. д. Каждый образец сразу же этикетировался, указывался район, область, условия местообитания, характер субстрата, дата, год и фамилия коллектора.
Собранные лишайники (листоватые, кустистые) сушили на воздухе. Лишайники сохнут хорошо и, как правило, не изменяют своей окраски. Лучше всего листоватые и кустистые лишайники собирать в пасмурную погоду, когда их слоевища эластичные и мягкие. Лишайники, собранные в сухую погоду, перед сушкой слегка смачивали водой для придания им эластичности.
Если субстрат (почва) влажен, то его просушивали на воздухе, чтобы лишайник не покрылся мицелием гриба и не испортился.
Эпифитные лишайники собирали вместе с перидермой, делая срезы как можно тоньше; в этикетке, кроме обычных сведений, указывали название древесной породы и на какой высоте взят образец лишайников. Высушенные и определенные лишайники перекладывали в чистовые гербарные пакеты. К верхней полосе конверта подклеивали этикетку, на которой указывали: название лишайника и семейство, местонахождение (область, район, деревня), время сбора, фамилия коллектора и лица, определившего образец.
3. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
3.1 Растительность Пинского лесхоза
Различие климатических, орографических и почвенно-гидрологических условий территории определяет зональные и местные изменения лесной растительности, которые, в свою очередь, находят отражение в лесорастительном районировании. Согласно существующему лесорастительному районированию территории Беларуси [18] территория Сошненского лесничества относится к Пинско-Припятскому лесному комплексу.
Распределение древесных пород характеризуется большим удельным весом сосны. Значительно распространены дубовые и черноольховые леса. Сравнительно большие площади заняты березой и незначительные елью. Встречаются ясеневые, грабовые, кленовые, липовые насаждения. По территории Пинского лесхоза проходит южная граница распространения ели. Из подлесочных пород здесь распространены: лещина, рябина, калина, крушина. В северной части встречается можжевельник.
3.2 Систематическая характеристика собранных видов
Отдел ЛИШАЙНИКИ – LICHENES
Класс СУМЧАТЫЕ ЛИШАЙНИКИ – ASCOLICHENES
Лишайники, у которых при половом размножении образуются сумки со спорами, размещающиеся в перитециях более или менее замкнутых плодоношениях (Pyrenolichenes) или апотециях – открытых плодовых телах (Discolichenes).
Порядок Стиктальные – Stictales
Слоевище гетеромерное, листоватое. Верхний и нижний коровой слой параплектенхимный, крупноклеточный. Апотеции с собственным и слоевищным краем, прикрепляются основанием или боком к лопастям слоевища. Сумки с 2-8 спорами. Споры поперечно-многоклеточные, иногда двухклеточные, бесцветные или темноватые [19].
Семейство Пельтигеровые Peltigeraceae
Род Пельтигера Peltigera Pers.
Пельтигера мягкая – Peltigera malacea (Ach.) Funk. (приложение 2, рисунок 6).
Таллом в виде широких, по краю округло-лопастных пластинок. Края лопастей лежат почти горизонтально либо слегка приподняты или загниты. Верхняя сторона зеленовато-коричневая, матовая, при смачивании зеленеющая, нижняя сторона розоватая. Жилки очень широкие, крупные, сливающиеся в почти сплошной войлок, коричневые, по краю светлые. Характерны крупные трещины на верхней стороне таллома и неясное жилкование по розоватому широкому краю снизу. Апотеции на вертикальных лопастях, плоские. На песчаных почвах и замшелых камнях в лесу, на лугах. Всегда на сухих местах.
Порядок Лецидеальные – Lecideales
Слоевище гетеромерное, корковое, листоватое, кустистое или других переходных типов.
Апотеции лецидеиновые или биаториновые, с хорошо выраженным собственным краем. Сумки с 1-8 спорами. Споры бесцветные и окрашенные, одноклеточные, поперечно-многоклеточные или муральные. Водоросли зеленые.
Семейство Кладониевые Cladoniaceae
Слоевище гетеромерное, однообразнонакипное или по переферии лопастное, чешуйчатое или листоватое, покрыто коровым слоем или без него, прикрепляется к субстрату ризинами или корнеподобными гаптерами, длительно сохраняется или со временем исчезает. На поверхности первичного слоевища развиваются подеции: простые, цилиндрические, палочковидные, шиловидные, кубковидные или разветвленные до кустистых, покрытые коровым слоем или без него, полые внутри или заполненные более или менее рыхлой тканью, голые или покрытые чешуйками (филлокладиями). Апотеции образуются на концах подециев или сбоку, иногда на чешуйках первичного слоевища, лецидеиновые или биаториновые. Парафизы большей частью не разветвленные. Сумки с 6-8 спорами. Споры бесцветные одноклеточные или поперечно-многоклеточные. Пикнидии размещены на подециях или чешуйках слоевища. Водоросли одноклеточные Coccomyxa, Pleurococcus.
Род Кладония – Cladonia Web.
Лишайник имеет первичный и вторичный таллом. Первичный таллом у немногих видов накипный, у большинства мелко- или реже крупночешуйчатый. У многих видов первичный таллом сохраняется на всем протяжении существования лишайника, у других быстро исчезает, и в этом случае сохраняется только вторичный таллом (подеции), развившийся на первичном. Подеции внутри полые и имеют разнообразную форму: палочковидные, сцифовидные (в виде бокальчика) или в виде кустиков с более или менее разветвленными округлыми ветвями. Сцифовидные подеции часто пролифелируют, т.е. из центра сцифы или по ее краям развиваются новые подеции. На подециях у многих видов бывают развиты филлокладии. Апотеции биаториновые, красные, бледно- или темно-коричневые. Развиваются на подециях. Один из самых больших родов кустистых лишайников.
Виды с кустистыми сильноразветвленными подециями
Кладония оленья – Cladonia rangiferina (L.) Web. (приложение 2, рисунок 7).
Первичный таллом отсутствует. Подеции сероватые или серовато-беловатые, до 20 см выс., сильноразветвленные, особенно в верхней части, с поникающими в одну сторону конечными веточками, верхушки которых обычно окрашены в темно-коричневый цвет. Образует густые дерновники. От KOH желтеет. Апотеции очень мелкие, коричневые. На песчаных почвах, на торфяниках, в светлых сосновых лесах, тундрах. Самая крупная среди кладоний с древовидно разветвленными подециями. На болотах иногда имеет розоватый оттенок и достигает 40 см. выс.
Кладония лесная – Cladonia sylvatica (L.) Hoffm. (приложение 2, рисунок 8).
Первичный таллом отсутствует. Подеции серовато- или желтовато-зеленые, до 10 см выс., сильноразветвленные и в местах ветвлений часто с отверстиями. Конечные веточки многочисленные, односторонне отогнутые вниз, с коричневатыми кончиками, от KOH слабо желтеют или совсем не изменяют окраски, жгуче-горьковатые на вкус. Апотеции коричневатые. На песчаных почвах, торфяниках, гнилых пнях, на открытых местах, в борах.
Кладония неприглаженная Cladonia impexa Harm. (приложение 2, рисунок 9).
Первичный таллом отсутствует. Подеции светло-желтоватые или зеленовато-серые, до 10 см выс. Основные стволики довольно толстые, в верхней части имеют вид густых, не закругленных кустиков. Конечные веточки растопырены во все стороны, но не загнуты вниз. Поверхность основных стволиков войлочная или бугорчатая. От КОН не изменяются. Апотеции редки, мелкие, коричневые. На песчаной почве в сосновых лесах, преимущественно на открытых местах, а также среди мхов на гнилых пнях.
Виды со сцифовидными разветвленными и неразветвленными подециями (наряду со сцифовидными у ряда видов могут присутствовать палочковидные подеции)
Кладония роговидная – Cladonia cornuta (L.) Schaer. (приложение 2, рисунок 10).
Первичный таллом из небольших чешуек, с возрастом исчезает. Подеции серовато- или зеленовато-коричневые, до 7-12 см выс. и 1-5 мм толщ., палочковидные, заостренные или сцифовидно расширенные, простые или слаборазветвленные, в верхней трети с мучнистым налетом из соредий, в нижней – с коровым слоем в виде бугорков. От KOH желтеют. Апотеции темно-коричневые. На перегнойной и песчаной почве, среди мхов, на гниющих пнях.
Кладония вздутая – Cladonia turgida (Ehrh.) Hoffm. (приложение 2, рисунок 11).
Первичный таллом из очень крупных, до 2,5 см дл., листовидно-лопастных чешуек, желтоватых или коричневато-зеленых сверху и беловатых снизу, сохраняется или с возрастом исчезает. Подеции бледно-зеленоватые, гладкие или местами потрескавшиеся, редко с филлокладиями, вздуто-цилиндрические, лучисто или вильчато разветвленные, 2-8 см и более выс., без сциф или сцифовидно расширенные. На дне сциф и в местах ветвлений отверстия. От KOH подеции не изменяются или слабо желтеют. Иногда подеции совсем не развиваются, и остается первичный таллом. Апотеции красновато-коричневые. На почве, на замшелых камнях, поваленных стволах, реже на пнях, в сосновых лесах и на открытых местах.
Кладония чешуйчатая – Cladonia squamosa (Scop.) Hoffm. (приложение 2, рисунок 12).
Первичный таллом из некрупных чешуек, обычно долго сохраняется, реже исчезает. Подеции серовато- или зеленовато-белые, кустистые, сцифовидно расширенные, пролиферирующие, с отверстиями на дне сциф и в местах ветвлений, до 7 см. выс., густо покрытые мелкими серовато-беловатыми чешуйками. От KOH не изменяются. Апотеции небольшие, коричневатые. На земле, замшелых скалах и гниющей древесине.
Кладония бесформенная Cladonia deformis Hoffm. (приложение 2, рисунок 13).
Первичный таллом из крупных широких чешуек, долго сохраняется. Подеции соломенно- или серо- желтые, вверху с тонкомучнистым соредиозным налетом, сцифовидно расширенные или бесформенные, простые, с неровным зубчатым или разорванным краем, изредка палочковидные, до 7 см выс. От KOH не изменяются. Апотеции ярко-красные или реже желтоватые. На почве, замшелых скалах, пнях, стволах
Виды с палочковидными и шиловидными неразветвленными или слаборазвитыми подециями
Кладония рогатолучистая Cladonia cornutoradiata (Coem.) Sandst. (приложение 2, рисунок 14).
Первичный таллом из мелких рассеченных чешуек, долго сохраняется. Подеции до 7-10 см выс., пепельно- или зеленовато-серые, покрытые мучнистым налетом из соредий, простые или чаще в верхней части роговидно разветвленные, шиловидные или с редкими скошенными узкими сцифами, лучисто пролиферирующими по краям. Окраска подециев от KOH не изменяется. Апотеции темно-коричневые. На почве, замшелых скалах, пнях.
Кладония вильчатая – Cladonia furcata (Huds.)Schrad. (приложение 2, рисунок 15).
Первичный таллом из мелких чешуек, обычно быстро исчезает. Подеции до 5-10 см выс., зеленовато-серые или коричневые, с коровым слоем, местами потрескавшимся, и с филлокладиями в нижней части, дихотомически разветвленные, неправильно цилиндрические, в местах ветвления продырявленные, на концах заостренные, всегда без сциф. От KOH слабо желтеют. Апотеции мелкие, коричневатые. На открытых местах и в светлых лесах на почве среди мхов, на замшелых камнях, на болотах.
Кладония шишконосная – Cladonia coniocraea (Flk.) Sandst. (приложение 2, рисунок 16).
Первичный таллом крупночешуйчатый, хорошо развит, долго сохраняется. Чешуйки оливково-зеленые, сильно изрезанные, до 0,5 см дл.. Подеции беловатые или беловато-зеленоватые, с мучнистым соредиозным налетом на поверхности, 0,3 – 3 см выс., простые, иногда в верхней части слегка разветвленные, шиловидные или тупороговидные, редко образуют очень узкие сцифы. От KOH подеции не изменяются или слабо желтеют. Апотеции темно-коричневые или рыжеватые, довольно редки. На основаниях стволов деревьев, замшелых пнях, старой древесине (заборы, крыши построек и т. д.).
Кладония бокальчатая – Cladonia pyxidata (L.)
Первичное слоевище в виде плотных изрезанных по краю чешуек, серых или зеленоватых сверху и белых снизу. Подеции до 3 см высоты, простые, образующие пролифицирующие сцифы. Поверхность подециев зеленовато-серая. Коровой слой хорошо развит при основании подециев, в верхней части и на внутренней части сциф кора в виде отдельных бугорков. Филлокладии развиваются, но не всегда. Апотеции коричневые. От KOH слоевище не изменяется в окраске или слабо желтеет, от Br и Pd оно краснеет. На почве по открытым солнцу полянам и склонам в сосновых и смешанных лесах, на разрушающейся древесине и у основания стволов, реже на кочках болот [4.].
Кладония дюймовая Cladonia uncialis (L.) (приложение 2, рисунок 17).
Первичное слоевище скоро исчезающее. Подеции до 8 см высоты, коротко дихотомически ветвящиеся, с отверстиями в местах ветвлений. Верхняя часть веточек подециев вертикально стоящая или поникающая, более темная, чем подеции, шиловидно заостренная или с 2-5 зубчиками. Поверхность подециев гладкая, несколько блестящая. Коровой слой хорошо выражен, сплошной или прерывистый. Филлокладии отсутствуют.
Апотеции светло-коричневые, мелкие. При действии КОН окраска слоевища не изменяется.
На песчаной почве по открытым солнцу полянам в сосновых лесах, на кочках по болоту.
На территории Белоруссии встречается повсеместно [1].
Род Стереокаулон Stereocaulon Schreb.
Кустистый лишайник с первичным и вторичным талломом. Первичный таллом зернистый, зернисто-бугорчатый или чешуйчатый, в виде корочки, сохраняется или быстро исчезает. Подеции простые, палочковидные или в виде сильноразветвленных кустиков, густо покрыты по всей поверхности филлокладиями разнообразной формы, беловатые, с розоватым или сероватым оттенком, редко темные. Кроме филлокладиев на поверхности подециев могут находиться цефалодии в виде головчато-бугорчатых выростов, темно- или светлоокрашенных. Апотеции коричневые или черные, биаториновые или лецидеиновые, расположены на верхушках главных стволиков (конечные) или на веточках (боковые).
Стереокаулон войлочный Stereocaulon tomentosum Fr. (приложение 2, рисунок 18).
Горизонтальное слоевище отсутствует. Псевдоподеции до 3-4 см высоты, 1-1,5 мм толщины, цилиндрические, в нижней части слабо, а вверху густо разветвленные, собраны в рыхлые дерновинки, которые слабо прикреплены к субстрату. Поверхность псевдоподециев беловато-серая, покрыта густым мохнатым войлоком, по ней неравномерно разбросаны филлокладии: снизу меньше, а сверху густо сидящие. Филлокладии голубовато-серые или грязновато-белые, чешуевидные, около 0,5 мм в диаметре, по краям зазубренные или рассеченные, более или менее сливающиеся или рассеянные.
Апотеции обчно многочисленны, конечные или боковые, до 0,5-1 мм в диаметре. Цефаллодии плохо различимые, скрытые в опушении, полушаровидные, 0,3-0,8 мм. На почве и замшелых камнях.
Порядок Леканоральные Lecanorales
Слоевище гетеромерное, накипное, листоватое или кустистое. Апотеции леканориновые, с хорошо развитым слоевищным краем, содержащим водоросли, сидячие или погруженные в слоевище. Сумки с 1-8 или многочисленными спорами. Споры одноклеточные, поперечно-многоклеточные или муральные, бесцветные или темные. Пикноконидии экзо- или эндобазидиальные.
Семейство Пармелиевые Parmeliaceae
Слоевище гетеромерное, чешуйчатое, листоватое или редко кустистое, дорсовентральное, прикрепляется к субстрату ризинами, нижней поверхностью слоевища или только центральной частью ее. Верхний коровой слой всегда присутствует и хорошо развит, нижний развит не всегда. Гонидиальный слой образован зелеными водорослями Trebouxia, сплошной или прерывистый, лежит под верхней корой, редко отдельные скопления водорослей встречаются у нижней коры. Сердцевина белая или желтая, иногда с довольно крупными полостями. Апотеции лекариновые, размещены по всему слоевищу или по краю лопастей, большей частью сидячие. Сумки с 8 (редко больше) спорами. Споры одноклеточные, бесцветные.
Род Гипогимния - Hypogymnia Nyl.
Таллом листоватый, в центре довольно плотно прикрепляется к субстрату участками нижней поверхности, без ризин, по краю с приподнимающимися или реже свисающими лопастями; лопасти часто вздутые. Сверху таллом серовато-зеленоватый, беловато-сероватый, желтовато-сероватый или коричневатый, снизу темный, от коричневато-черного до черного, к краям может быть немного светлее, или, наоборот, нижняя поверхность может быть развита лучше верхней, и тогда она выступает в виде черной каймы по краям лопастей. Поверхность таллома снизу складчатая или морщинистая, иногда с округлыми или овальными отверстиями. На концах лопастей или по всей их поверхности часто развиваются соредии. Апотеции сидячие или на ножках, развиваются редко. Растут преимущественно на стволах и ветвях древесных пород, реже на обработанной древесине, замшелых скалах, почве.
Гипогимния вздутая H. physodes (L.) Nyl. (приложение 2, рисунок 19).
Таллом листоватый, очень разнообразный по форме (розетковидный, полурозетковидный или неопределенной формы), с тесно сближенными или налегающими друг на друга лопастями 1-5 см дл. И 1-6 мм шир., слегка выпуклыми, разветвленными, слегка расширенными, вздутыми, внутри полыми. По краю лопастей очень часто расположены белые мучнисто зернистые соредии. Сверху таллом серовато-зеленоватый, желтовато- серовато-зеленоватый или голубовато-зеленовато-серый, гладкий или морщинистый-складчатый, снизу черный или темно-коричневый, морщинистый, без отверстий. Апотеции сидячие или на коротких ножках, коричневые, встречаются редко. Преимущественно на стволах и ветвях хвойных и лиственных пород, реже на других субстратах. Часто растет с другими видам гипогимний. Один из самых обычных лесных видов [20].
Род Пармелия – Parmelia Ach.
Таллом листоватый, разрезано-лопастной, в виде крупных розеток, прикрепленный к субстрату ризинами, реже свободный. Лопасти разнообразные: узкие или широкие, сильно- или маловетвистые, плоские или выпуклые, тесно сомкнутые или раздельные. Верхняя сторона от беловато-сероватой и желтоватой до коричневатой и черной, матовая или блестящая; нижняя – от белой или светло-коричневой до черной. Хорошо развиты ризины, простые или ветвистые. Часто развиваются соредии и изидии разнообразной формы. Апотеции леканориновые, сидячие или на ножках, развиваются по всей поверхности талломов, часто более обильно в центре. Эпифиты на коре деревьев, реже на замшелых почвах, на обнаженной древесине.
Пармелия бороздчатая – Parmelia sulcata Tayl. (приложение 2, рисунок 20).
Таллом неправильно-розетковидный, 5-15 см в диам. Лопасти 3-4 мм шир. и 5-20 мм дл., выемчатые, тесно собранные или немного расходящиеся, на концах тупые. Верхняя сторона таллома голубовато- или зеленовато-серая, сетчато-морщинистая, с соредиями; нижняя – черная, густо покрыта до концов лопастей черными, простыми или ветвящимися ризинами. Апотеции до 2 см в диам., коричневые, с вогнутым диском, редки. Таллом от KOH желтеет. Очень полиморфный вид. На стволах и ветвях лиственных, реже хвойных пород, на обработанной древесине и каменистом субстрате, обычно на хорошо освещенных местах [21].
Род Цетрария – Cetraria Ach.
Таллом листоватый, слабо прикрепляющийся к субстрату или кустистый, из вертикально стоящих лопастей, собранных вместе, с хорошо выраженной верхней и нижней сторонами. Верхняя сторона беловато- или зеленовато-сероватая, желтоватая либо коричневатая; нижняя – от светлой до темной, часто тех же оттенков, что и верхняя, с немногочисленными ризинами или без них. Апотеции лекариновые, развиваются на концах лопастей, сидячие или на короткой ножке. На ветвях и стволах деревьев, на почве среди мхов и других видов лишайников, на больших камнях, валунах.
Цетрария исландская («исландский мох») – Cetraria islandica (L.) Ach. (приложение 2, рисунок 21).
Таллом имеет вид беловато- или зеленовато-коричневых кустиков, состоящих из плоских, желобчато-завернутых или почти трубчатых лопастей до 10 см выс. и 0,5-4 см шир. Внизу лопасти красноватые или с отдельными красными пятнами. Края лопастей обычно с ресничками, иногда на вершинах лопастей они отсутствуют. Соредии и изидии редки. Нижняя сторона лопастей немного светлее верхней и обычно с макулами. Апотеции редки, развиваются на краях или концах расширенных лопастей, того же цвета, что и таллом, до 1,5 см в диам. От KOH таллом желтеет. Очень полиморфный вид, варьирующий в основном по форме и величине лопастей, наличию или отсутствию соредий и изидий, окраске. Растет на почве в сосновых лесах, на болотах среди мхов. Встречается как целыми дерновинами, так и отдельными экземплярами среди других лишайников и мхов.
Цетрария заборная – Cetraria saepincola (Ehrh.) Ach.
Слоевище кожисто-листоватое, плотно прилегающие к субстрату, часто розетковидное, лопастное. Слоевищные лопасти короткие, неправильно рассеченные. Верхняя поверхность слоевища зеленовато-коричневая, коричневая или оливково-коричневая, матовая, чуть морщинистая, не растрескавшаяся. Соредии и изидии отсутствуют. Нижняя сторона слоевища коричневатая или более светлая, со слабым жилкованием и рассеченными длинными, иногда ветвистыми белыми ризинами. Верхняя кора темная, коричневатая или оливковая, параплектенхиматическая. Гонидиальный слой непрерывный, компактный. Сердцевинный слой беловатый, рыхлый. Нижняя кора аналогична верхней.
Апотеции лекариновые, встречаются в изобилии, размещаются на концах лопастей. Диск апотеция красно-коричневый, плоский или слегка выпуклый, без налета, чуть глянцеватый. Слоевищный край тонкий, гладкий или зазубренный. Эпитеций бледно-коричневый. Гимений хорошо выражен. Сумки широкобулавовидные. Споры одноклеточные, эллипсовидные, бесцветные. Пикнидии темные, размещены по краю лопастей. Пикноконидии простые, в середине перетянутые.
Слоевище от реактивов, применяемых для диагностики лишайников, окраски не меняет.
Встречается на стволах и ветвях хвойных и лиственных пород, на веточках кустарника, иногда на обработанной древесине. Особенно обильно на молодых ветвях берез.
Семейство Усниевые – Usneaceae
Слоевище кустистое, листовато-кустистое или листоватое, многократно разветвленное, прикрепляется к субстрату ризоидами или ясно выраженным основанием, свисающее, стелющееся или торчащее, различной величины и цвета у разных видов. Слоевищные лопасти (веточки) округлые в сечении, радиального строения или дорсовентральные, верхняя и нижняя поверхности отличаются по цвету. Кора образована из гиф, более или менее параллельно уложенных к оси лопасти и образующих параплектенхимную ткань. У некоторых видов образуется механическая ткань, лежащая под корой. Под корой размещается сердцевина, образованная сплетением толстостенных гиф. Гонидиальный слой образован водорослями Cystococcus. Апотеции леканориновые, блюдцевидные, довольно крупные. Сумки с 8 спорами. Споры одно- или двухклеточные, чаще бесцветные. Соредии имеют различное строение.
Род Эверния – Evernia Ach.
Таллом кустистый, мягкий, из округлых или плоских, дихотомически ветвящихся лопастей; прикреплен к субстрату гомфом. Центрального тяжа нет. Сердцевинная ткань рыхлая, паутинистая. Апотеции редки, и если есть, то довольно крупные (до 10мм в диам.). Растет на коре деревьев, обнаженной древесине, почве.
Эверния шелушащаяся – Evernia furfuracea (L.) Mann. (приложение 2, рисунок 22).
Таллом кустистый, состоит из лопастей с желобчато-загнутыми на нижнюю сторону краями. Верхняя сторона светло-зеленовато-серая до темно-серой, с многочисленными изидиями; нижняя – синевато-черная или голубовато-серая; у молодых экземпляров нижняя сторона розоватая. Лопасти правильно дихотомически ветвятся. На коре деревьев, обработанной гнилой древесине.
Род Рамалина – Ramalina Ach.
Таллом кустистый, прямостоячий или повисающий, большей частью жесткий, более или менее блестящий. Лопасти таллома лентовидно-сплюснутые, реже цилиндрически-округлые, дихотомически разветвленные; коровой слой лопастей на разрезе выглядит зазубренным, что связано с особенностями анатомического строения сердцевины. Апотеции лекариновые, боковые или конечные, по краю без ресничек. На коре деревьев. Почве, камнях.
Рамалина ясеневая – Ramalina fraxinea (L.) (приложение 2, рисунок 23).
Таллом кустистый, повисающий вниз или распростертый по субстрату, длинный (до 20 см), очень жесткий. Лопасти сплюснутые, довольно широкие (иногда до 1 см), с сетчато-складчатой поверхностью, серовато-зеленоватые. Апотеции развиваются очень часто и сидят на коротких ножках по бокам лопастей. Диск грязновато-розовый. На коре лиственных, реже хвойных деревьев.
Порядок Калоплакальные Caloplacales
Слоевище гетеромерное, корковое, листоватое или кустистое, включает водоросли Cystococcus. Апотеции лекариновые или лецидеиновые, содержат хризофеновую кислоту и от KOH окрашиваются в вишнево-красный цвет. Споры 2-4 клеточные (реже одноклеточные) с округлыми или линзовидными клеточными просветами, полярные, клетки спор соединяются между собой каналом.
Семейство Телосхистовые Teloschistaceae
Род Ксантория – Xantoria Th. Fr.
Таллом листоватый, в виде округлых розеток или небольших оранжево-желтых подушечек. Поверхность таллома от KOH моментально окрашивается в вино-красный цвет. Апотеции лекариновые, в центре или по краю таллома, сидячие или прижатые, обычно обильные.
Ксантория постенная (стенная золотнянка) – Xantoria parietina (L.) (приложение 2, рисунок 24).
Таллом свыше 3 см в диам., в виде правильных оранжево-желтых розеток, состоящих из крупных, широких, округлых по краю лопастей. На концах лопасти выемчато-изрезанные. В центре таллома многочисленные апотеции, диск которых обычно окрашен ярче таллома. Эпифит, обитающий на коре деревьев, растет также на обработанной древесине, особенно часто на заборах и стенах построек.
Семейство Фисциевые – Physciaceae
Слоевище листоватое или кустистое, гетеромерное с верхним и нижним коровым слоем, прикрепляется к субстрату ризоидами или гифами подслоевища. Апотеции леканориновые или лецидеиновые, округлые, сидячие или чуть приподняты над субстратом. Сумки с 8 спорами. Споры 2-4 клеточные, редко мурально-многоклеточные, коричневые, с утолщенными стенками. Водоросли Cystococcus. Пикноконидии эндобазидиальные, прямые.
Род Фисция – Physcia Fr.
Таллом листоватый, в виде правильных розеток, реже в виде чешуйчатых корочек, плотно прирастающих к субстрату с помощью многочисленных ризоидов, обычно заметных при рассматривании таллома сверху в виде мелких ресничек по краю лопастей. Верхняя поверхность обычно плотная, ровная, гладкая, без морщинок и неровностей, б. ч. Сероватая или коричневатая, часто с беловатым или сизым налетом. Апотеции лекариновые. На коре деревьев или на камнях.
Фисция сизая – Physcia caesia (Hoffm.) Hampe. (приложение 2, рисунок 25).
Таллом в виде листовидных розеток, 2-3 см в диам., сверху коричневато-сероватый, снизу светлый, с рассеянными черными ризоидами, которые могут развиваться в виде оторочки по краям лопастей. Лопасти вильчато разветвленные, выпуклые и только на концах плоские, плотно прижатые одна к другой. Таллом от KOH желтеет. На концах боковых отростков лопастей и в центре таллома развиваются крупные головчатые свинцово- или беловато-серые сорали. Апотеции редки. Диск их красновато-коричневый, с беловатым налетом, окружен толстым слоевищным краем. На камнях, преимущественно на известняках, реже на обнаженной древесине, на коре лиственных пород и почве.
Фисция звездчатая Physcia stellaris (L.) Nyl. (приложение 2, рисунок 26).
Таллом в виде плотно приросших к субстрату розеток, сверху беловатых или сизовато-серых, снизу светлых, с густыми серыми ризоидами. Лопасти узкие, вытянутые, на концах округло-выемчатые. Сверху таллом от КОН желтеет. Апотеции почти всегда развиваются, с темно-коричневым диском, иногда покрытым беловатым налетом. На коре лиственных деревьев, особенно часто на тополе, осине, реже на коре хвойных и на камнях [4].
Таблица 1 – Систематический список объектов исследования
| № | Таксон | Местообитание (фитоценоз) |
|
Семейство Пельтигеровые – Peltigeraceae Род Пельтигера – Peltigera Pers. |
||
| 1 |
Пельтигера мягкая – Peltigera malacea (Ach.) Funk. |
Сосняк мшистый (Pinetum pleurozisum) |
|
Семейство Кладониевые – Cladoniaceae Род Кладония – Cladonia Web. |
||
| 2 |
Кладония оленья Cladonia rangiferina (L.) Web. |
Сосняк мшистый (Pinetum pleurozisum) |
| 3 |
Кладония лесная – Cladonia sylvatica (L.) Hoffm. |
Сосняк мшистый (Pinetum pleurozisum) |
| 4 |
Кладония неприглаженная – Cladonia impexa (L.) |
Сосняк лишайниковый (Pinetum cladinosum) |
| 5 |
Кладония роговидная – Cladonia cornuta (L.) Schaer. |
Сосняк мшистый (Pinetum pleurozisum) |
| 6 |
Кладония вздутая – Cladonia turgida (Ehrh.) Hoffm. |
Сосняк лишайниковый (Pinetum cladinosum) |
| 7 |
Кладония чешуйчатая – Cladonia squamosa (Scop.) Hoffm. |
Сосняк лишайниковый (Pinetum cladinosum) |
| 8 |
Кладония бесформенная – Cladonia deformis Hoffm. |
Сосняк мшистый (Pinetum pleurozisum) |
| 9 |
Кладония рогатолучистая – Cladonia cornutoradiata (Coem.) Sandst. |
Сосняк мшистый (Pinetum pleurozisum) |
| 10 |
Кладония вильчатая – Cladonia furcata (Huds.)Schrad. |
Сосняк мшистый (Pinetum pleurozisum) |
| 11 |
Кладония шишконосная – Cladonia coniocraea (Flk.) Sandst. |
Сосняк лишайниковый (Pinetum cladinosum) |
| 12 |
Кладония бокальчатая – Cladonia pyxidata (L.) |
Сосняк лишайниковый (Pinetum cladinosum) |
| 13 |
Кладония дюймовая Cladonia uncialis (L.) |
Сосняк лишайниковый (Pinetum cladinosum) |
|
Род Стереокаулон Stereocaulon Schreb. |
||
| 14 |
Стереокаулон войлочный – Stereocaulon tomentosum Fr. |
Сосняк лишайниковый (Pinetum cladinosum) |
|
Семейство Пармелиевые – Parmeliaceae Род Гипогимния - Hypogymnia Nyl. |
||
| 15 |
Гипогимния вздутая – Hypogymnia physodes (L.) Nyl. |
Сосняк мшистый (Pinetum pleurozisum) |
|
Род Пармелия – Parmelia Ach. |
||
| 16 |
Пармелия бороздчатая – Parmelia sulcata Tayl. |
Сосняк мшистый (Pinetum pleurozisum) |
|
Род Цетрария – Cetraria Ach. |
||
| 17 |
Цетрария исландская («исландский мох») Cetraria islandica (L.) Ach. |
Сосняк лишайниковый (Pinetum cladinosum) |
| 18 |
Цетрария заборная – Cetraria saepincola (Ehrh.) Ach. |
Сосняк мшистый (Pinetum pleurozisum) |
|
Семейство Усниевые – Usneaceae Род Эверния – Evernia Ach. |
||
| 19 |
Эверния шелушащаяся – Evernia furfuracea (L.) Mann. |
Сосняк мшистый (Pinetum pleurozisum) |
|
Род Рамалина – Ramalina Ach. |
||
| 20 |
Рамалина ясеневая – Ramalina fraxinea (L.) |
Сосняк мшистый (Pinetum pleurozisum) |
|
Семейство Телосхистовые - Teloschistaceae Род Ксантория – Xantoria Th. Fr. |
||
| 21 |
Ксантория постенная (стенная золотнянка) – Xantoria parietina (L.) |
Сосняк мшистый (Pinetum pleurozisum) |
|
Семейство Фисциевые – Physciaceae Род Фисция - Physcia Fr. |
||
| 22 |
Фисция сизая – Physcia caesia (Hoffm.) Hampe. |
Сосняк мшистый (Pinetum pleurozisum) |
| 23 |
Фисция звездчатая Physcia stellaris (Ach.). |
Сосняк мшистый (Pinetum pleurozisum) |
Таблица 2 – Распределение объектов исследования по жизненным формам и экологическим группам.
| № | Название лишайника | Жизненная форма | Экологическая группа |
| 1 |
Пельтигера мягкая – Peltigera malacea (Ach.) Funk. |
Листоватый | Эпигейный (напочвенный) |
| 2 |
Кладония оленья Cladonia rangiferina (L.) Web. |
Кустистый | Эпигейный (напочвенный) |
| 3 |
Кладония лесная – Cladonia sylvatica (L.) Hoffm. |
Кустистый | Эпигейный (напочвенный) |
| 4 |
Кладония неприглаженная – Cladonia impexa (L.) |
Кустистый | Эпигейный (напочвенный) |
| 5 |
Кладония роговидная – Cladonia cornuta (L.) Schaer. |
Кустистый | Эпигейный (напочвенный) |
| 6 |
Кладония вздутая – Cladonia turgida (Ehrh.) Hoffm. |
Кустистый | Эпигейный (напочвенный) |
| 7 |
Кладония чешуйчатая – Cladonia squamosa (Scop.) Hoffm. |
Кустистый | Эпигейный (напочвенный) |
| 8 |
Кладония бесформенная – Cladonia deformis Hoffm. |
Кустистый | Эпигейный (напочвенный) |
| 9 |
Кладония рогатолучистая – Cladonia cornutoradiata (Coem.) Sandst. |
Кустистый | Эпигейный (напочвенный) |
| 10 |
Кладония вильчатая – Cladonia furcata (Huds.)Schrad. |
Кустистый | Эпигейный (напочвенный) |
| 11 |
Кладония шишконосная – Cladonia coniocraea (Flk.) Sandst. |
Кустистый | Эпигейный (напочвенный) |
| 12 |
Кладония бокальчатая – Cladonia pyxidata (L.) |
Кустистый | Эпигейный (напочвенный) |
| 13 |
Кладония дюймовая Cladonia uncialis (L.) |
Кустистый | Эпигейный (напочвенный) |
| 14 |
Стереокаулон войлочный – Stereocaulon tomentosum Fr. |
Кустистый | Эпигейный (напочвенный) |
| 15 |
Гипогимния вздутая –Hypogymnia physodes (L.) Nyl. |
Листоватый | Эпифитный |
| 16 |
Пармелия бороздчатая – Parmelia sulcata Tayl. |
Листоватый | Эпифитный |
| 17 |
Цетрария исландская («исландский мох») Cetraria islandica (L.) Ach. |
Кустистый | Эпигейный (напочвенный) |
| 18 |
Цетрария заборная – Cetraria saepincola (Ehrh.) Ach. |
Кустистый | Эпифитный |
| 19 |
Эверния шелушащаяся – Evernia furfuracea (L.) Mann. |
Кустистый | Эпифитный |
| 20 |
Рамалина ясеневая – Ramalina fraxinea (L.) |
Кустистый | Эпифитный |
| 21 |
Ксантория постенная (стенная золотнянка) – Xantoria parietina (L.) |
Листоватый | Эпифитный |
| 22 |
Фисция сизая – Physcia caesia (Hoffm.) Hampe. |
Листоватый | Эпифитный |
| 23 |
Фисция звездчатая Physcia stellaris (Ach.). |
Листоватый | Эпифитный |
ЗАКЛЮЧЕНИЕ
В результате проведенной работы, на территории Сошненского лесничества Пинского лесхоза было собрано и определено 23 вида лишайников, которые входят в состав 6 семейств:
Семейство Пельтигеровые Peltigeraceae
Семейство Кладониевые Cladoniacea
Семейство Пармелиевые Parmeliacea
Семейство Усниевые – Usneaceae
Семейство Телосхистовые Teloschistaceae
Семейство Фисциевые – Physciaceae
Наиболее часто встречаемыми оказались представители семейства Кладониевые – Cladoniaceae, их численность составила 13 видов:
Кладония оленья Cladonia rangiferina (L.) Web.
Кладония лесная Cladonia sylvatica (L.) Hoffm.
Кладония роговидная – Cladonia cornuta (L.) Schaer.
Кладония вздутая – Cladonia turgida (Ehrh.) Hoffm.
Кладония чешуйчатая – Cladonia squamosa (Scop.) Hoffm.
Кладония бесформенная – Cladonia deformis Hoffm.
Кладония рогатолучистая Cladonia cornutoradiata (Coem.) Sandst.
Кладония вильчатая Cladonia furcata (Huds.)Schrad.
Кладония шишконосная Cladonia coniocraea (Flk.) Sandst.
Кладония бокальчатая Cladonia pyxidata (L.)
Кладония дюймовая Cladonia uncialis (L.)
Виды этого семейства развиваются на песчаной почве сухих сосновых лесов, которыми преобладает территория Сошненского лесничества, а также на бесплодных, хорошо освещаемых местах, на опушках леса, у основания стволов деревьев. Представители рода кладония широко распространены по всему земному шару.
Мы убедились в том, что лишайники обладают способностью расти на самых различных субстратах, в самых неблагоприятных условиях, что даёт им возможность конкурировать с быстрорастущими растениями и мхами. На изучаемой территории мы обнаружили 2 экологические группы лишайников: эпигейных – 65,22% (15 видов), эпифитных 34,78% (8 видов), эпилитных – 0%.
Собранные лишайники представлены следующими жизненными формами: кустистые – 73,91% (17 видов), листоватые – 26,09% (6 видов), накипные – 0%.
Разнообразие жизненных форм и преобладание среди них кустистых лишайников говорит о том, что на исследуемой территории чистый атмосферный воздух.
Редко встречающимися видами на данной территории являются Пельтигера мягкая – Peltigera malacea (Ach.) Funk, Цетрария заборная – Cetraria saepincola (Ehrh.) Ach., Рамалина ясеневая – Ramalina fraxinea (L.), Эверния шелушащаяся – Evernia furfuracea (L.) Mann.
Перспективами нашей работы является уточнение видового состава лихенофлоры, выявление новых видов и изучение практического использования лишайников.
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1. Горбач Н. В. Лишайники Белоруссии. Определитель. «Наука и техника». Мн., 1973 (3)
2. Солдатенкова Ю. П. Малый практикум по ботанике. Лишайники. М., Изд-во Моск. ун-та, 1977 (3)
3. Жизньрастений. Т.3. /Ред. М.М.Голлербах/, Водоросли. Лишайники. М., Просвещение, 1977, 487 с.
4. Водоросли, лишайники и мохообразные СССР. Отв. Ред. М.В. Горленко. М., «Мысль», 1978. (67, 68-70, 70-83, 84)
5. Яковлев Г. П., Челомбитько В. А. Ботаника: Учеб. для фармац. институтов и фармац. фак. мед. вузов/Под ред. И. В. Грушвицкого. – М.: Высш. шк., 1990. – 367 с, ил. (143)
6. Пчелкин А. В. Популярная лихенология. – М. МГСЮН, 36 с. (5)
7. Лемеза Н. А., Шуканов А. С. Малый практикум по низшим растениям: Учеб. пособие. Мн.: Унiверсiтэцкае, 1994. – 288 с.
8. Бигон М., Харпер Дж., Таунсенд К. Экология. Особи, популяции и сообщества: В 2-х т. Т. 1.: Пер. с англ. – М.: Мир, 1989. – 667 с, ил. (650)
9. Стрельская О. Я. Низшие растения. Систематика: [Учеб. пособие для пед. ин-тов по биол. и геогр. спец.] / Под ред. Н. А. Дорожкина. – Мн.: Выш. шк., 1985. 240 с., ил.
10. Ботанический атлас: Под общ. ред. чл. – корр. АН СССР Б. К. Шишкина. М. – Л., Сельхоиздат, 1963.
11. Бязров Л.Г. Лишайники в экологическом мониторинге. – М.: Научный мир, 2002. 336с.
12. Великанов Л.Л., Сидорова И.И., Успенская Г.Д. Полевая практика по экологии грибов и лишайников. - Изд-во МГУ, 1980, 112 с.
13. Горышина Т. К. Экология растений: Учеб. пособие. – М.: Высш. школа, 1979. – 368 с., ил. (91)
14. Блукет Н. А., Емцев В. Т. Ботаника с основами физиологии растений и микробиологии. Изд. 2-е, перераб. и доп. М., «Колос», 1974. – 560 с. , ил. (300).
15. Стекольников Л.И., Мухор В.И. Целебные кладовые природы. Мн.:Ураджай, 1981. – 271с. (53)
16. Пчелкин А. В., Слепов В. Б. Использование водорослей и лишайников в экологическом мониторинге и биоиндикационных исследованиях. – М., Московская городская станция юных натуралистов, 2004, 20 (2)
17. Пчелкин А. В., Боголюбов А. С. Методы лихеноиндикации загрязнений окружающей среды. Методическое пособие. М., Экосистема, 1997, 25 с. (4)
18. Юркевич И.Д., Гельтман В.С. География, типология и районирование лесной растительности Белоруссии. Минск, 1965
19. Комарницкий Н.А., Лишайники, бактерии и актиномицеты. М., Изд-во «Высшая школа», 1960. – 294 с.
20. Н.С.Голубкова. Определитель лишайников средней полосы Европейской части СССР. М.-Л.: "Наука", 1966.
21. Определитель лишайников России. Вып.6. Алекториевые, Пармелиевые, Стереокаулоновые. Под. ред. Н.С.Голубковой. С.-П.: "Наука", 1996.
© 2010 Интернет База Рефератов