
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Дипломная работа: Сравнительный анализ структуры наследственной компоненты подверженности к бронхиальной астме и туберкулезу по генам ферментов метаболизма ксенобиотиков
Дипломная работа: Сравнительный анализ структуры наследственной компоненты подверженности к бронхиальной астме и туберкулезу по генам ферментов метаболизма ксенобиотиков
РОССИЙСКАЯ АКАДЕМИЯ МЕДИЦИНСКИХ НАУК СИБИРСКОЕ ОТДЕЛЕНИЕ ТОМСКИЙ НАУЧНЫЙ ЦЕНТР ГОСУДАРСТВЕННОЕ УЧРЕЖДЕНИЕ НАУЧНО-ИССЛЕДОВАТЕЛЬСКИЙ ИНСТИТУТ МЕДИЦИНСКОЙ ГЕНЕТИКИ
ГОСУДАРСТВЕННОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ
СИБИРСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ ФЕДЕРАЛЬНОГО АГЕНТСТВА ПО ЗДРАВООХРАНЕНИЮ И СОЦИАЛЬНОМУ РАЗВИТИЮ
На правах рукописи
БРАГИНА
ЕЛЕНА ЮРЬЕВНА
СРАВНИТЕЛЬНЫЙ АНАЛИЗ СТРУКТУРЫ НАСЛЕДСТВЕННОЙ КОМПОНЕНТЫ ПОДВЕРЖЕННОСТИ К БРОНХИАЛЬНОЙ АСТМЕ И ТУБЕРКУЛЕЗУ ПО ГЕНАМ ФЕРМЕНТОВ МЕТАБОЛИЗМА КСЕНОБИОТИКОВ
03.00.15. – генетика
Диссертация
на соискание ученой степени
кандидата биологических наук
Научный руководитель:
академик РАМН,
профессор В. П. Пузырев
ТОМСК-2005
ОГЛАВЛЕНИЕ
Список сокращений 4
Введение 6
Глава 1. Обзор литературы 12
1.1. Ферментативная система биотрансформации ксенобиотиков 12
1.1.1. Cемейства ферментов I и II фаз метаболизма 12
1.1.2. Свойства ферментов метаболизма ксенобиотиков 14
1.1.3. Генетический полиморфизм ферментативной системы метаболизма ксенобиотиков 17
1.2. Молекулярно-генетические аспекты мультифакториальных заболеваний (бронхиальная астма и туберкулез) 21
1.3. Полиморфизм генов ферментов биотрансформации ксенобиотиков и патология 37
Глава 2. Материал и методы исследования 48
2.1. Характеристика обследованных групп населения 48
2.1.1. Характеристика группы больных туберкулезом 48
2.1.2. Характеристика группы больных бронхиальной астмой 50
2.2. Характеристика методов исследования 52
2.2.1. Клинико-лабораторные методы исследования 52
2.2.2. Молекулярно-генетические методы исследования 54
2.2.3. Статистические методы анализа 57
Глава 3. Результаты и обсуждение 60
3.1. Полиморфизм генов глутатионовых S-трансфераз (GSTT1, GSTM1, GSTP1) и цитохромов Р450 (CYP2E1, CYP2C19) у жителей г. Томска 60
3.2. Оценка роли полиморфизма генов ферментов метаболизма ксенобиотиков в развитии бронхиальной астмы и туберкулеза 65
3.2.1. Ассоциация полиморфных вариантов генов GSTT1, GSTM1, GSTP1, CYP2E1 и CYP2C19 с атопической бронхиальной астмой 65
3.2.2. Ассоциация полиморфизма генов ферментов метаболизма ксенобиотиков с туберкулезом 70
3.2.3. Сравнительный анализ роли полиморфных вариантов генов ферментов метаболизма ксенобиотиков в детерминации бронхиальной астмы и туберкулеза 76
3.3. Анализ ассоциаций генов ферментов метаболизма ксенобиотиков с бронхиальной астмой и туберкулезом на семейном материале 78
3.4. Оценка связи комбинаций генотипов генов ферментов биотрансформации ксенобиотиков с туберкулезом и бронхиальной астмой 81
3.5. Связь полиморфизма генов ферментов метаболизма ксенобиотиков с изменчивостью количественных признаков у больных бронхиальной астмой и туберкулезом 85
Заключение 101
Выводы 107
Литература 109
СПИСОК СОКРАЩЕНИЙ
95% CI – 95% доверительный интервал;
CYP – гены цитохрома Р450;
GST – глутатион S-трансфераза;
GSTT1 (θ1) – глутатион S-трансфераза тета 1;
GSTT1+ - гомо- и гетерозиготы гена GSTT1;
GSTМ1 (μ1) - глутатион S-трансфераза мю 1;
GSTМ1+ - гомо- и гетерозиготы гена GSTМ1;
GSTР1 (π1) - глутатион S-трансфераза пи 1;
HLA главный комплекс гистосовместимости человека;
Ig – иммуноглобулины;
IL1B ген интерлейкина 1 В;
IL1RN ген антагониста рецептора к интерлейкину 1;
INF-γ – гамма интерферон;
mEH микросомальная эпоксигидролаза;
NAT2 – ген N-ацетилтрансферазы;
NRAMP1 (NRAMP1) – ген макрофагального белка (макрофагальный белок), ассоциированного с естественной резистентностью;
OR (Odds ratio) – отношение шансов;
P450 – цитохромы Р450;
S.D. – стандартное отклонение;
S.E. – стандартная ошибка;
TDT (Transmission/Disequilibrium Test) – тест на неравновесие по сцеплению;
TNFА ген фактора некроза опухолей;
VDR ген рецептора к витамину D;
АБП – антибактериальные препараты;
АЛТ аланинаминотрасфераза;
АСТ аспартатаминотрансфераза;
БА – бронхиальная астма;
БГР (BHR) – бронхиальная гиперреактивность;
ИЛ – интерлейкин (ы);
МБТ (M. tuberculosis) микобактерия туберкулеза;
ОМЛ – острая миелоидная лейкемия;
ОФВ1 (FEV1) – объем форсированного выдоха за первую секунду;
ПСВ (PEF)– пиковая скорость выдоха;
РС20 – наличие бронхиальной гиперреактивности, установленное с помощью ингаляционного провокационного теста с метахолином;
РЛ – рак легкого;
РРП – рак ротовой полости;
РХФ – равновесие Харди-Вайнберга;
САП – скарификационные аллергопробы;
ТБ – туберкулез;
ФВД – функции внешнего дыхания;
ФЖЕЛ (FVC) – форсированная жизненная емкость;
ФМК/ФБК – ферменты метаболизма/биотрансформации ксенобиотиков.
ВВЕДЕНИЕ
Актуальность проблемы.
Генетика широко распространенных болезней человека является активно развивающейся областью исследований. Однако темп накопления сведений о конкретных генах, участвующих в их возникновении и развитии существенно уступает известным на сегодня знаниям по генетике моногенных (менделевских) болезней. Еще более скромные успехи отмечены в изучении генетических основ подверженности к инфекционным заболеваниям. В последнем случае преобладают исследования, касающиеся изучения генетических характеристик возбудителей болезней, их геномов в формировании восприимчивости (устойчивости) человека к конкретной инфекции и клинического полиморфизма болезни. Наряду с этим направлением – изучение генома самого человека, контактирующего с инфекцией, заболевшего или сохранившего здоровье - становится важной областью генетических исследований [Пузырев и др., 2002; Frodshem, Hill, 2004]. Заметим, что отечественным генетиком А.С. Серебровским (1939) было высказано положение, обозначенное им как противоречие «единства бесконечного числа признаков и конечного числа генов», нашедшее, спустя более полувека, развитие в геномных исследованиях человека и обсуждение проектов «Феном человека» [Freimer, Sabatti, 2003] и «Феном мыши» [Paigen, Eppig, 2000]. «Важное различие между геномом и феномом состоит в том, что в то время как геном ограничен (приблизительно 3 млрд. пар оснований у человека), феном – нет (его предел зависит от того, как далеко мы хотим двигаться)» - эта мысль, сформулированная K. Paigen и J.T. Eppig (2000) тождественна положению А.С. Серебровского (1939). Подмеченное сходство взглядов классика генетики XX века и современных исследователей генома человека на гено-фенотипические взаимоотношения [Пузырев, 2001] является, по нашему мнению, обоснованием перспективности высказываемых и ранее гипотез о том, что клинически различные группы (нозологии) заболеваний человека могут контролироваться общим набором генов подверженности [Becker et al., 1998].
С позиции изучения вклада «общих» генов в развитие различных болезней особую актуальность приобретает исследование системы генов метаболизма ксенобиотиков, поскольку ферментами этой системы осуществляется метаболизм не только большинства разнообразных по химической структуре экзогенных молекул, но и многочисленных эндогенных веществ, например, медиаторов воспаления. Система ферментов метаболизма ксенобиотиков представляет собой сформировавшийся в процессе эволюции механизм адаптации организма к воздействию токсичных экзогенных и эндогенных веществ. Предполагается, что различия в скорости деградации различных субстратов ферментами метаболизма могут лежать в основе неодинаковой восприимчивости к ряду заболеваний. Изучению участия генов этой системы в развитии онкопатологии, эндометриоза, бронхиальной астмы, хронической обструктивной болезни легких, инфекционных заболеваний посвящены многие работы отечественных и зарубежных авторов [Lin et al., 1998; Иващенко и др., 2001; Ляхович и др., 2000, 2002; Delfino et al., 2000; Вавилин и др., 2002; Rollinson et al., 2003; Бикмаева и др., 2004]. Очевидно, что генетические различия в регуляции, экспрессии и активности генов ферментов биотрансформации ксенобиотиков являются решающими факторами в развитии болезни и позволяют рассматривать ее как важное звено в этиологии и патогенезе этих заболеваний.
Особое внимание исследователей привлекает участие ферментативной системы метаболизма в биотрансформации лекарственных препаратов [Nebert, 1997]. Изучение полиморфизма генов этой системы в различных популяциях, обусловливающего существование индивидуальных особенностей метаболизма лекарственных препаратов, проявляющихся различиями в эффективности терапии и наличием многообразных побочных эффектов медикаментозной нагрузки, являются достаточно перспективными в практическом применении.
Представляется перспективным проведение сравнительного анализа участия белков ферментов метаболизма ксенобиотиков в возникновении и развитии заболеваний, которые с одной стороны, часто сочетаются друг с другом у одного индивидуума (синтропии), с другой – редко или совсем не встречаются вместе (дистропии).
Туберкулез (ТБ) и бронхиальная астма (БА), являющиеся частой патологией народонаселения, по-видимому, относятся к дистропным заболеваниям. Так, эпидемиологическая парадигма свидетельствует о том, что риск развития атопической БА и ее различных клинических проявлений в течение жизни намного ниже у индивидов, перенесших ТБ в детском возрасте [Von Hertzen et al., 1999, Shirakawa et al., 1997]. Тем не менее, показано, что при БА и ТБ имеет место общая генетическая основа (гены системы HLA, интерлейкинов и их рецепторных антагонистов и др.), обусловленная функциональной значимостью продуктов экспрессии этих генов в инфекционно-аллергическом процессе [Sandford et al., 1996; Greenwod et al., 2000; Bellamy, 2000; Sengler et al., 2002].
Таким образом, изучение роли полиморфных вариантов генов системы метаболизма в развитии БА и ТБ актуально и предполагает исследование их связи с клиническими особенностями течения заболеваний для понимания механизмов взаимодействия в процессе реализации наследственной информации на уровне целостного организма.
Цель работы: Провести сравнительный анализ значения полиморфизма генов ферментов метаболизма ксенобиотиков в развитии бронхиальной астмы и туберкулеза легких, оценить их роль в формировании клинических проявлений данных заболеваний у жителей города Томска.
Задачи исследования:
1. Изучить распространенность частот полиморфных вариантов генов ферментов метаболизма ксенобиотиков (CYP2C19, CYP2E1, GSTT1, GSTM1 и GSTP1) в выборке здоровых индивидов.
2. Оценить связь полиморфизмов исследуемых генов с атопической бронхиальной астмой и туберкулезом легких.
3. Изучить связь полиморфных вариантов, включенных в исследование генов, с клиническими особенностями течения бронхиальной астмы и туберкулеза легких, а также с патогенетически значимыми для этих заболеваний качественными и количественными признаками.
4. Провести сравнительный анализ роли полиморфных вариантов генов системы метаболизма ксенобиотиков в развитии бронхиальной астмы и туберкулеза.
Научная новизна:
Получены новые знания о роли генов ферментов биотрансформации ксенобиотиков (GSTT1, GSTM1, GSTP1, CYP2E1, CYP2C19) в развитии бронхиальной астмы и туберкулеза легких у жителей города Томска. Впервые проведена сравнительная оценка значимости исследуемых полиморфных вариантов генов системы метаболизма в развитии бронхолегочных патологий (на примере бронхиальной астмы и туберкулеза). Выявлены ассоциации полиморфизма генов GSTM1 (делеция) и CYP2E1 (7632T>A) с развитием бронхиальной астмы, а GSTP1 (313A>G) – с туберкулезом. Изучено влияние полиморфных вариантов генов системы метаболизма на развитие различных клинических особенностей течения заболеваний. Впервые проведена сравнительная оценка относительного риска в зависимости от комбинаций генотипов исследуемых генов для развития бронхиальной астмы и туберкулеза. Установлена роль генов глутатионовых S-трансфераз (GSTT1, GSTM1, GSTP1) и цитохромов Р450 (CYP2C19, CYP2E1) в детерминации изменчивости количественных, патогенетически значимых для заболеваний признаков. Показана связь полиморфного варианта 313A>G гена GSTP1 с изменчивостью уровня аланинаминотрансферазы у больных туберкулезом легких во время лечения антимикобактериальными препаратами.
Практическая значимость:
Полученные результаты исследования могут быть положены в основу разработки скрининговых программ по выявлению лиц с повышенным риском развития бронхиальной астмы и туберкулеза. Сведения о связи полиморфных вариантов генов ферментов метаболизма ксенобиотиков с изменчивостью показателей печеночной функции могут быть учтены при проведении профилактических мероприятий с целью предотвращения проявлений гепатотоксичности во время противотуберкулезной терапии. Материалы работы могут быть использованы в учебно-методическом процессе на биологических и медицинских факультетах ВУЗов. Полученная информация о полиморфизме генов ферментов биотрансформации ксенобиотиков у русских жителей города Томска может быть использована при проведении генетико-эпидемиологических исследований широко распространенных заболеваний.
Положения, выносимые на защиту:
1. Генетическими маркерами подверженности к бронхиальной астме могут быть генотип Т/А (полиморфизм 7632Т>А) гена CYP2E1 и «нулевой» генотип делеционного полиморфизма гена GSTM1.
2. У жителей города Томска генотип G/G гена GSTP1 (полиморфизм 313A>G) снижает риск развития туберкулеза.
3. Фактором генетической предрасположенности к бронхиальной астме является «нулевой» генотип гена GSTM1 как в сочетании с генотипом GSTT1+, так и в комбинации с гетерозиготным генотипом гена CYP2E1 (полиморфизм 7632Т>А).
4. «Нулевой» генотип гена GSTM1 и генотип *1/*1 гена CYP2C19 оказывают влияние на формирование клинических фенотипов бронхиальной астмы, определяющихся такими показателями как: уровень общего иммуноглобулина Е в сыворотке крови и форсированная жизненная емкость легких.
5. Изменчивость признаков, характеризующих особенности клинического течения туберкулеза (уровень эритроцитов и аланинаминотрансферазы), определяется полиморфными вариантами генов CYP2C19 (681G>A) и GSTP1 (313A>G) системы метаболизма ксенобиотиков.
Апробация работы:
Основные результаты исследования по теме диссертационной работы доложены и обсуждены на межлабораторных научных семинарах ГУ НИИ медицинской генетики ТНЦ СО РАМН (Томск, 2002, 2003); VI, VII научных конференциях «Генетика человека и патология» (Томск, 2002, 2004); IV Международном конгрессе молодых ученых «Науки о человеке» (Томск, 2003); V съезде Российского общества медицинских генетиков (Уфа, 2005).
ГЛАВА 1. ОБЗОР ЛИТЕРАТУРЫ
1.1. Ферментативная система биотрансформации ксенобиотиков
1.1.1. Семейства ферментов I и II фазы метаболизма
В процессах метаболизма различных по химическому составу ксенобиотиков, в том числе лекарственных препаратов и ряда эндогенных субстратов, выделяют две фазы [Urs, 1997]. Цитохромы Р450, флавинсодержащие монооксигеназы, эстеразы, амидазы, альдегиддегидрогеназы и др. относят к ферментам I-й фазы биотрансформации, которые участвуют в реакциях окисления и восстановления, а также гидролиза молекул ксенобиотика [Gonzalez, 1993]. Ведущая роль в окислении многих ксенобиотиков, а также важнейших для жизнедеятельности эндогенных соединений, таких как стероидные гормоны, витамины, жирные и желчные кислоты, простагландины, лейкотриены, биогенные амины, ретиноиды и др. принадлежит цитохрому Р450 [Ляхович, Цырлов, 1981; Waxman, Azaroff, 1992]. В ходе ферментативных реакций I-й фазы биотрансформации (фаза активации) образуются водорастворимые соединения. В дальнейшем эти соединения могут подвергаться конъюгации с эндогенными соединениями, восстановлению или гидролизу с помощью ферментов II-й фазы (фаза детоксикации), а затем выведению из организма. Ко второй фазе метаболизма принадлежат ферменты конъюгации – глутатион S-трансферазы (GST), конъюгирующие главным образом электрофильные соединения с глутатионом, УДФ-глюкуронозилтрансферазы (UDPGT), катализирующие реакции конъюгации молекул ксенобиотика или его метаболита с глюкуроновой кислотой [Morgenstern, DePierre, 1985], N-ацетил- (NAT), сульфо- (ST) -трансферазы, эпоксидгидролазы (EH), гидролизующие эпоксиды и др. [Sipes, Gandolfi, 1986].
В реакции II-й фазы метаболизма ксенобиотики могут вступать не только после метаболизма в реакциях I-й фазы, но и напрямую, а впоследствии подвергаться или не подвергаться окислению ферментами цитохрома Р450 [Saito et al., 1986], а результатом метаболизма может быть как уменьшение, так и усиление токсичных свойств субстрата. На рис. 1 представлены возможные комбинации взаимодействия двух фаз биотрансформации.
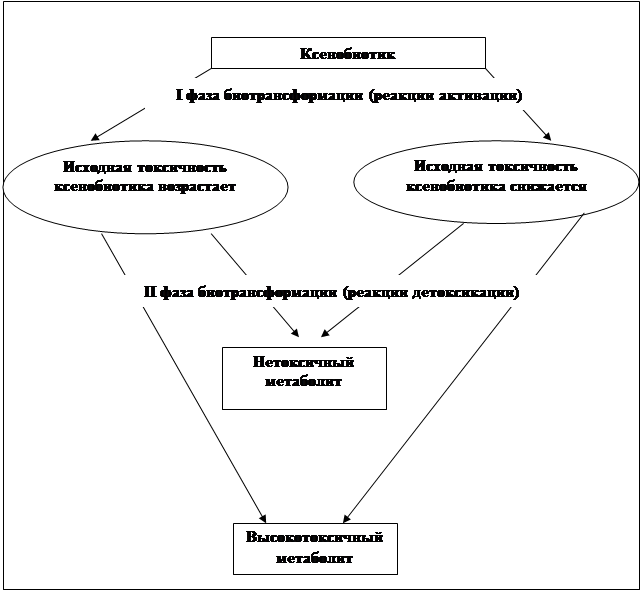
Рис.1. Изменение токсичных свойств ксенобиотиков в ходе реакциий I-й и II-й фаз биотрансформации.
Наиболее благоприятным исходом из них будет вариант, когда изначально токсичные свойства ксенобиотика снижаются под воздействием ферментов I и II фазы, а высокая активность различных цитохромов Р450 в сочетании с низкой активностью ферментов II-й фазы биотрансформации является наиболее неблагоприятной и приводит к увеличению риска развития некоторых заболеваний [Guengerich, 1988].
1.1.2. Свойства ферментов метаболизма ксенобиотиков
Цитохром Р450 является уникальным по своим свойствам гемопротеидом, обеспечивающим внедрение активированного кислорода непосредственно в молекулу субстрата. В общей сложности известно о 107 генах цитохромов Р450 в геноме человека, из них 59 индивидуальных цитохромов Р450 и 48 псевдогенов [Ingelman-Sundberg, 2004]. На сегодняшний день для большинства цитохромов установлена функциональная значимость. Цитохромы Р450 семейств 1-3 ответственны в большинстве случаев (70-80% из всех ферментов I-й фазы биотрансформации) за метаболизм используемых в клинической практике лекарственных препаратов [Ingelman-Sundberg, 2004; Evans, Relling, 1999; Bertz, Granneman, 1997]. Члены семейства CYP1, 2, 3, 4 – ответственны за метаболизм чужеродных соединений, а CYP11, CYP17, CYP19, CYP21 вовлечены в метаболизм стероидов и желчных кислот [Ioannides, Lewis, 2004; Lewis et al., 2004; Rifkind et al., 1995]. Часть цитохромов Р450 окисляют жирорастворимые витамины, некоторые вовлечены в метаболизм жирных кислот и эйкозаноидов.
Для многих цитохромов Р450 описаны высокоспецифичные субстраты. Однако одной из особенностей как цитохрома Р450, так и его индивидуальных форм является способность к метаболизму большого спектра субстратов. Поэтому изоформы цитохрома Р450 перекрываются в своей субстратной специфичности, и даже высокоспецифичные субстраты могут подвергаться метаболизму многими из них [Райс, Гуляева, 2003]. Интересно, что наряду с селективными субстратами существуют и такие, в метаболизме которых участвуют многие формы цитохрома Р450. Классическим примером такого субстрата является лекарственное средство антипирин, который метаболизируют CYP1A1, 2C8, 2C9, 2C18, 2B6, 3A4, 2D6, 2A6, 2C19 и 2Е1 [Engel et al., 1996].
Глутатион S-трансферазы – мультигенное семейство соответствующих ферментов, которое участвует в метаболизме большого числа электрофильных соединений путем их конъюгации с глутатионом, а также в биотрансформации некоторых эндогенных соединений (гормонов, липидов, простагландинов, лейкотриенов) [Morgenstern, DePierre, 1985; Кулинский, 1999; Hayes, Strange, 1999]. К настоящему времени известно, что у млекопитающих различают 6 подклассов глутатион S-трансфераз: 5 семейств цитоплазматической (альфа (α), мю (μ), тэта (θ), пи (π) и зета (Z)) и одно семейство микросомальной GST [Eaton, Bammler, 1999] . Синтез глутатионовых S-трансфераз контролируется различными генами, в которых выявлены полиморфизмы, оказывающие существенное влияние на их функции. Известно, что функциональная GST является димером [Beckett, Hayes, 1993].
Цитохром Р450 первоначально был обнаружен в печени, а затем и в других органах. Изучение внепеченочной экспрессии позволило сказать о тканеспецифичности цитохромов Р450. Тканеспецифичная экспрессия различных изоформ цитохрома Р450 определяет особенности протекающих монооксигеназных реакций и отражает адаптацию этой универсальной ферментной системы к структурно-функциональной организации той или иной системы организма. Так, высокая экспрессия цитохрома Р450 в гепатоцитах обеспечивает наиболее активное участие этого органа в биотрансформации ксенобиотиков. В печени ферменты метаболизма ксенобиотиков представлены максимально, а затем по убыванию следуют почки, легкие, кишечник, головной мозг и другие органы. В надпочечниках и половых железах в основном экспрессированы изоформы, участвующие в биосинтезе стероидных гормонов, в почках - изоформы, участвующие в биотрансформации ксенобиотиков и витамина Д и т.д. [Ingelman-Sundberg et al., 1995; Haehner et al., 1996].
На протяжении дыхательного тракта экспрессируются как цитохромы P450, так и ферменты второй фазы биотрансформации. Так в различных сегментах легких обнаружены ферменты семейств CYP1, 2, 3 и 4 [Wheeler, Guenthner, 1991; Raunio et al., 1995]. Из ферментов второй фазы наиболее представлены по всей протяженности респираторного тракта NAT1, NAT2, а также GSTμ1, GSTμ3 и GSTπ1. Необходимо отметить, что глутатионовые S-трансферазы p класса составляют более чем 90% от общей GST-активности в эпителиальных клетках легких человека [Frayer et al., 1986] .
Таким образом, знания об экспрессии генов ферментов метаболизма в различных органах и тканях, а также выявление их субстратной специфичности создают возможность объяснения тканеспецифичного метаболизма ксенобиотиков [Ravindranath, 1998]. Однако для этого необходимо изучение специфичного взаимодействия ферментов I-й и II-й фазы в метаболизме различных по химическому составу эндогенных и экзогенных ксенобиотиков, в том числе и лекарственных препаратов, определение их активности и генотипирования полиморфных генов [Pelkonen, Raunio, 1997; Nebert et al., 2003].
Одним из важных свойств системы цитохрома Р450 является индукция – активация транскрипции гена в присутствии субстрата [Ляхович, Цырлов; 1981]. Ранее предполагалось, что ксенобиотики сами являются факторами регуляции собственного метаболизма, однако впоследствии были показаны генетические механизмы процесса индукции [Poland et al., 1973]. Cпособность к индукции характерна для многих генов ферментов метаболизма ксенобиотиков семейств цитохрома Р450 [Honkakoski, Negishi, 2000] и имеет для организма приспособительное значение к меняющимся условиям химического окружения [Denison, Whitlock, 1995], в некоторых случаях достаточно довольно низких концентраций ксенобиотиков-индукторов, чтобы вызвать сильный ответ [Whitlock, Gelboin, 1974; Surry et al., 2000].
Некоторые ксенобиотики оказывают противоположный индукции эффект – ингибируют активность цитохромов Р450, что происходит вследствие образования реактивного метаболита, который ковалентно фиксируется в активном центре фермента. Показано ингибирование активности ферментов некоторыми лекарствами, например, изониазидом [Wen et al., 2002]. В случае, когда несколько ксенобиотиков метаболизируются одним и тем же ферментом семейства цитохрома Р450, они являются конкурентными ингибиторами друг для друга.
1.1.3. Генетический полиморфизм ферментативной системы метаболизма ксенобиотиков
Молекулярные механизмы полиморфизма генов ферментов метаболизма ксенобиотиков обусловлены следующим:
a) Нуклеотидные различия в кодирующем регионе гена приводят к замене аминокислоты и изменению в деятельности фермента или связывания субстрата (например, CYP2D6).
б) Делеции в кодирующем регионе приводят к отсутствию фермента или недостаточному синтезу белка (например, CYP2A6, CYP2D6 и GSTM1).
в) Полиморфизмы в некодирующей области затрагивают элементы транскрипционного контроля, вовлеченные в экспрессию и индукцию фермента (например, CYP1A1).
г) Изменения в сигнале полиаденилирования изменяет количество фермента (например, NAT1).
д) Генная амплификация повышает количество фермента (например, CYP2D6).
е) Сложные взаимодействия полиморфных генов и/или их ферментативных продуктов (например, более высокая активность CYP1A1 и 1A2 у лиц с GSTM1-дефицитом, вероятно из-за большего бионакопления компонентов индукции) [Bartsch et al., 2000].
С феноменом генетического полиморфизма ферментов, участвующих в биотрансформации ксенобиотиков впервые столкнулись фармакологи, и это явление обусловливает значительные межиндивидуальные различия в метаболизме – до 104 [Guengerich, 2003]. По причине существования многочисленных данных с использованием различных обозначений аллелей генов цитохромов Р450 в настоящее время выработана единая классификация, рекомендованная к применению для исследователей [Nelson et al., 1996].
У человека подкласс GSTμ кодируется генами, локализованными на хромосоме 1 в области 1р13.3 и включает пять тандемно расположенных генов: GSTM1, GSTM2, GSTM3, GSTM4 и GSTM5 [Афанасьева, Спицин, 1990]. Для гена GSTM1 установлены две мутации: точковая замена, не имеющая функциональных проявлений [De Long et al., 1988], и протяженная делеция гена (10 т.п.н.), которая возникла в результате неравного кроссинговера между двумя гомологичными последовательностями, фланкирующими ген GSTM1, проявляющаяся отсутствием белка [Seidegard, 1988]. GSTM1*A и GSTM1*B кодируют GSTM1A и GSTM1B ферменты, которые функционально идентичны и различаются только по одной аминокислоте. GSTM1A содержит лизин в позиции 172, а GSTM1B – аспарагинин в этом же положении [Hatagima, Strange, 2000].
Ген GSTT1 картирован на хромосоме 22 (локус 22q11.2). Его полиморфизм обусловлен наличием двух аллелей: функционально активного GSTT1*1 и неактивного, так называемого «нулевого» (GSTT1*0). Аллель GSTT1*0 соответствует частичной или полной делеции, приводящей к снижению активности белка [Pemble et al., 1994].
Ген GSTP1 локализован на хромосоме 11 (11q13) и преимущественно экспрессируется в альвеолярных клетках, альвеолярных макрофагах, бронхиолах и плаценте. Для гена GSTP1 описаны две точковые мутации: замена аденина на гуанин в 313 положении первичной последовательности GSTP1, проявляющейся заменой изолейцина 105 на валин (Ile105Val) в 5 экзоне, и замена С341Т, проявляющейся заменой аланина 114 на валин (Ala114Val) в 6 экзоне [Board et al., 1989]. При мутации 105Val в 7 раз увеличивается каталитическая активность фермента по отношению к полициклическим ароматическим соединениям, но в 3 раза снижена активность по отношению к 1-хлор-2,4-динитробензену [Watson et al., 1998].
К настоящему моменту описаны девять аллелей гена CYP2C19, два активных аллеля CYP2C19*1A (wt1) и CYP2C19*1B (wt2) и семь дефектных аллелей CYP2C19*2A (m1A), 2C19*2B (m1B), 2C19*3 (m2), 2C19*4 (m3), 2C19*5A (m4 или TRP433), 2C19*5B, и 2C19*6 (m5) [Romkes et al., 1991; Richardson et al., 1995; Ibenau et al., 1998]. Основной генетический дефект, найденный у «медленных» метаболизеров (S)-мефенитоина – точечная замена G на A в пятом экзоне в положении 681 гена CYP2C19 (CYP2C19*2), приводящая к аберрантному сайту сплайсинга. Образующаяся мРНК не содержит первые 40 оснований пятого экзона, что нарушает рамку считывания, и приводит к образованию стоп-кодона. В печени индивидуумов, гомозиготных по этому дефекту, обнаруживается лишь аберрантно сплайсированная РНК. Таким образом, сплайсинг проходит исключительно с использованием сайта, возникшего в результате мутации [Крынецкий, 1996]. Этот полиморфизм является важным в отношении метаболизма лекарственных препаратов, связанный с нарушением способности цитохрома Р450 метаболизировать антиэпилептический препарат (S)-мефенитоин, а также омепразол, прогуанил, некоторые барбитураты и др. Кроме того показана еще одна точечная замена G→A в положении 636 в четвертом экзоне гена CYP2C19 (CYP2C19*3), приводящая к продукции укороченного белка [Ibenau et al., 1999; Xie et al., 1999; Yang et al., 2004; Schwab et al., 2004].
Ген CYP2E1 локализован на хромосоме 10q24.3-qter и состоит из 11413 п.н. и содержит 9 экзонов, кодирующих продукт из 493 аминокислот [Kolble, 1993]. Для гена CYP2E1 (табл. 1) наиболее часто рассматриваются тесно сцепленные полиморфизмы по рестрикционным эндонуклеазам PstI/RsaI (мутантный аллель CYP2E1*5B), локализованные в 5’-фланкируещем регионе гена [Hayashi et al., 1991; Watanabe et al., 1994;], при которых мутантный аллель способствует повышенной транскрипционной и ферментативной активности, а также DraI полиморфизм (мутантный аллель CYP2E1*6), расположенный в 6 интроне [Uematsu et al., 1991], для редкого аллеля которого показаны мутации, влияющие на экспрессию гена и каталитическую активность соответствующего белка [Hu et al., 1997].
Таблица 1
Номенклатура аллелей CYP2E1 гена (составлена по данным сайта http://www/imm.ki.se/CYPalleles)
| Аллель | Белок | Однонуклеотидные замены |
Эндонуклеаза рестрикции |
|
CYP2E1*1A CYP2E1*1B CYP2E1*1C CYP2E1*1D CYP2E1*2 CYP2E1*3 CYP2E1*4 CYP2E1*5A CYP2E1*5B CYP2E1*6 CYP2E1*7A CYP2E1*7B CYP2E1*7C |
CYP2E1.1 CYP2E1.1 CYP2E1.1 CYP2E1.1 CYP2E1.2 CYP2E1.3 CYP2E1.4 CYP2E1.1 CYP2E1.1 CYP2E1.1 CYP2E1.1 CYP2E1.1 |
- 9893C>G 6 тандемов 8 тандемов 1132G>A 10023G >A 4768G>A -1293G>C -1053C>T 7632T>A -1293G>C -1053C>T 7632T>A -333T>A -71G>T;-333T>A -333T>A;-352A>G |
TaqI DraI, XbaI PstI RsaI DraI PstI RsaI DraI |
Таким образом, качественный состав и количественные соотношения изоформ ферментов метаболизма ксенобиотиков могут меняться под воздействием непосредственно самих же ксенобиотиков на организм. В зависимости от структуры исходного субстрата может происходить либо его биоактивация и увеличение токсичности, либо обезвреживание ксенобиотика. В результате ингибирования, индукции и генетического полиморфизма ферментов метаболизма ксенобиотиков может возникать дефицит или очень высокая активность отдельных изоформ и, как следствие, иметь место нежелательные для организма последствия: дисбаланс процессов биотрансформации ксенобиотиков, приводящий к развитию патологического состояния организма, а также снижение терапевтической активности лекарственных препаратов и всевозможные проявления побочных эффектов от их терапевтического действия.
1.2. Молекулярно-генетические аспекты мультифакториальных заболеваний (бронхиальная астма и туберкулез)
Развитие подавляющего большинства мультифакториальных заболеваний (МФЗ) происходит при сочетанном влиянии разнообразных факторов. МФЗ представляют группу болезней, развитие которых определяется неблагоприятным сочетанием полиморфных вариантов генов, контролирующих возникновение и патогенез заболевания в совокупности с определенными воздействиями факторов среды. Для МФЗ характерен ряд особенностей, которые с одной стороны, позволяют рассматривать эту группу патологий как модель изучения комплекса специфичных генов и экзогенных факторов, которые, взаимодействуя между собой, формируют норму реакции устойчивости человека к среде обитания [Гинтер, 2001; Бочков и др., 1984], а с другой - значительно осложняют обобщение данных для установления истинных генов подверженности сложнонаследуемых заболеваний. Например, существенное увеличение распространенности многих полигенных заболеваний (астма и связанные с атопией патологические состояния, туберкулез и др.) нельзя объяснить изменениями в генетической структуре за прошедшие десятилетия. Вероятно, что существующие генетические факторы, взаимодействующие с изменившимися условиями окружающей среды (снижение числа инфекционных болезней, повсеместная иммунизация, особенности питания и др.) вызывают повышенную восприимчивость популяции к вышеперечисленным заболеваниям [Organov, Maslennikova, 1999; Sengler et al., 2002]. Это пример того, как воздействие факторов внешней среды может значительно изменить положение порога подверженности к МФЗ [Фогель, Мотульски, 1990]. Кроме того, необходимо учитывать наличие сочетаний индивидуальных для каждой отдельно взятой популяции аллельных вариантов генов предрасположенности к заболеванию, что отражают различающиеся результаты анализа ассоциаций с МФЗ. Тем не менее, установление генов предрасположенности и изучение их совместной работы, выявление особенностей взаимодействия с факторами негенетической природы в развитии МФЗ, для которых пожизненный риск оценивается в западных популяциях порядка 60%, вызывает естественное стремление исследователей к пониманию механизмов нормальной и патологической реализации генетической информации [Пузырев, 2003].
Бронхиальная астма (БА) широко распространенное хроническое заболевание дыхательных путей, поражающее в России от 3 до 12 %, а в некоторых промышленно-развитых регионах эти цифры достигают 30 % [Научно-практическая программа «Бронхиальная астма у детей: диагностика, лечение и профилактика», 2004], а также порядка 5 миллионов детей и 10 миллионов взрослых в Западных странах [Schwartz et al., 2004]. Кроме того, отмечено повышение уровня числа больных, требующих госпитализации, а также рост показателей смертности от астмы. Несмотря на явные успехи в области выявления и лечения данной патологии, распространенность и тяжесть заболевания значительно увеличиваются за последние десятилетия.
Драматическое увеличение распространенности и тяжести астмы на протяжении последних 20 лет, особенно в ряде промышленных регионов предполагает, что ухудшающиеся условия окружающей среды играют далеко не последнюю роль в развитии и прогрессировании данной патологии. Отмеченное влияние ряда факторов, например, возраст, раса, социально-экономический статус, хотя и предполагает их участие в риске развития БА, но все-таки особую роль в этиологии и патогенезе заболевания отводят влиянию аллергенов, курению, профессиональным химическим агентам, загрязнителям воздуха, вирусам и иммунизации против конкретного инфекционного заболевания.
В 90% случаев выявления больных бронхиальной астмой присутствует атопия как генетически детерминированная способность организма к выработке повышенного IgE в ответ на воздействие аллергенов окружающей среды. Через IgE-опосредованный механизм целый ряд клеточных элементов: гистиоциты (тучные клетки), макрофаги, лимфоциты, эпителиальные и эндотелиальные клетки независимо друг от друга или совместно принимают участие в воспалении дыхательных путей, тем самым, осуществляя иммунный ответ организма на внедрение антигена. Воспалительная природа заболевания проявляется в морфологических изменениях стенки бронхов - дисфункции ресничек мерцательного эпителия, деструкции эпителиальных клеток, инфильтрации клеточными элементами, дезорганизации основного вещества, гиперплазии и гипертрофии слизистых и бокаловидных клеток. Длительное течение воспалительного процесса приводит к необратимым морфофункциональным изменениям в виде резкого утолщения базальной мембраны, нарушения микроциркуляции и склероза стенки бронха. Ключевой особенностью астмы является состояние бронхиальной гиперреактивности, свидетельствующее о повышенном бронхоконстрикторном ответе на различные физико-химические факторы, включая не только аллергены, к которым сенсибилизирован индивид, но и специфические стимулы, например, холодный воздух и физическая нагрузка [Гриппи, 1997]. Формирование гиперреактивности связывают с перестройкой дыхательных путей, обусловленной хроническим аллергическим воспалением, сопровождающейся сужением стенок, повышением васкуляризации, гипертрофией и гиперплазией гладкой мускулатуры бронхов. В результате чего происходят изменения нейрональной регуляции и повышение сократимости гладких мышц дыхательных путей. Как и атопия, неспецифическая гиперреактивность являются одними из универсальных признаков астмы: чем выше эти показатели, тем тяжелее протекает процесс. Однако распространенность бронхиальной гиперреактивности значительно выше, чем БА.
На протяжении более чем столетней истории вопроса наследования БА обсуждались различные модели моногенные (аутосомно-рецессивная и доминантная), полигенные, сцепленные с половыми хромосомами [Huang, Marsh, 1993; Чучалин, 1999]. В ходе исследований стало понятно, что сложные механизмы наследования астмы (как и атопии) не могут быть объяснены простой (моногенной) моделью, а проявление клинических симптомов болезни является результатом действия средовых факторов на предрасположенных индивидуумов [Anderson, Cookson, 1999].
Для оценки генетического вклада в этиологию и патогенез БА были предприняты массовые близнецовые исследования в Швеции, Финляндии, Норвегии, Дании, США и Австралии, показавшие оценку наследуемости от 15 до 75 %, что подтвердило предположение о генетической основе заболевания [Edfors-Lubs, 1971; Duffy et al., 1990; Nieminen et al., 1991; Lichtenstein, Svatengren, 1997; Laitinen et al., 1998; Skadhauge et al., 1999].
Большинство современных исследователей рассматривают генетическую компоненту заболевания БА как полигенную систему с аддитивным эффектом отдельных генов, каждый из которых в отдельности не способен, либо крайне редко способен вызвать болезнь [Holgate et al., 1995; LeSouef, 1997]. Таким образом, БА, как и многие распространенные заболевания в популяции, рассматривается как полигенная болезнь с наследственной предрасположенностью или как мультифакториальная болезнь. Для астмы, как и для остальных заболеваний этой группы характерны следующие признаки, сформулированные в 1969 году C.O. Carter: а) относительно высокая частота болезни в популяции и в то же время значительная семейная подверженность; б) наличие патогенетических и ассоциированных маркеров предрасположения; в) хроническое течение и наличие форм, образующих непрерывный ряд проявлений от ярко выраженных до субклинических; г) более раннее начало заболевания и утяжеление клинических симптомов в нисходящих поколениях семьи; д) относительно невысокая (в сравнении с моногенными болезнями) конкордантность по заболеванию у монозиготных близнецов; е) повышенный риск повторного рождения предрасположенных к болезни детей с появлением каждого последующего пораженного болезнью ребенка; ж) однотипность проявлений болезни у больного ребенка и ближайших родственников, что отражает коэффициент наследуемости, превышающий 50–60%; з) несоответствие закономерностей наследования болезни простым менделевским моделям (доминантное, рецессивное и др.) [Carter, 1996].
Таким образом, достижения в области исследования важнейших механизмов развития астмы позволили выработать концепцию патогенеза БА, согласно которой в основе клинических проявлений болезни лежит атопия, которая, как известно, характеризуется значительным вкладом наследственных факторов. А тщательная оценка эпидемиологии астмы позволяет определить экологические факторы риска БА.
Существует мнение, что контакт с бактериальными и вирусными инфекциями в раннем детстве является защитным фактором к дальнейшему развитию атопического заболевания в более поздней жизни. Еще в 1989 г. Strachan заметил, что распространение сенной лихорадки среди взрослых находится в обратной связи с размером семьи и даже более того – с наличием братьев и сестер [Strachan, 1989]. В связи с чем была выдвинута гипотеза, предполагающая, что инфекции в раннем детстве оказывают защитный эффект против развития в дальнейшем аллергии, получившая в последующем название «гигиенической гипотезы». С момента этого наблюдения выполнено много исследований, посвященных изучению связи между инфекциями, перенесенными в раннем периоде жизни и последующим развитием атопических заболеваний [Noguchi et al., 1998; Heinzmann et al., 2000]. Воссоединение Германии в 1990 г. способствовало уникальной возможности изучать распространение астмы в генетически схожих популяциях, но в условиях воздействия различных факторов окружающей среды, в том числе инфекции. Несмотря на то, что дети из бывшей Восточной Германии чаще болели инфекциями верхних дыхательных путей по сравнению с Западной Германией, развитие астмы в этих двух популяциях имело обратную зависимость [von Mutius et al., 1994]. В контексте «гигиенической гипотезы» интересны исследования, в которых показано, что дети, выросшие на ферме в тесном контакте с сельскохозяйственными и домашними животными, реже имели сенсибилизацию к пыльцевым и другим атопическим аллергенам в сравнении с детьми, выросшими в другой среде. Эти результаты указывают на то, что окружающая среда, характеризующаяся высоким содержанием бактерий, может действительно защищать от развития аллергии, по крайней мере, если субъект в раннем возрасте находился в такой среде. Основным механизмом данного защитного действия является способность эндотоксинов, содержащихся в бактериально загрязненной домашней пыли, стимулировать Th1-иммунитет [Ильина, 2001].
На сегодняшний день показано сцепление БА и ее клинических проявлений со многими хромосомными регионами. Изучение кандидатных генов показало сцепление с атопией и бронхиальной гиперреактивностью по многим локусам, но наибольшая важность показана для регионов 5q, 6p, 11q, 12q, 13q, 14q, 16p, и именно для этих локусов получены воспроизводимые результаты (табл. 2).
Таблица 2
Гены-кандидаты бронхиальной астмы и связанных с ней клинических фенотипов
| Локализация | Молекула | SNP/мутация |
Связанный фенотип |
Литературный источник |
| 1 | 2 | 3 | 4 | 5 |
| 1р32 | Гистамин-N-метил-трансфераза | С314Т(Thr105Ile) | Астма | Yan et al., 2000 |
| 1p13.3 | GSTM1 | del |
Астма Атопия |
Ляхович и др., 2000; Вавилин и др., 2002; Zhang et al., 2004 |
| 2q14 | IL1A | G/T at +4845 | Астма | Adjers et al., 2004 |
| 3p21 | СCR5 | CC5-∆32 | Астма | Hall et al., 1999 |
| 5q22-q24 | СYP1A1 | Аллель Val | Астма | Вавилин и др., 2002 |
| 5q31-34 | IL-4 | C-590T |
Астма/ Общий IgE/ Специфический IgE/ Атопический дерматит |
Rosenwasser et al., 1995; Walley et al., 1996; Noguchi et al., 1998; Kawashima et al., 1998; Burchard et al., 1999 |
| С+33Т | Астма/ Общий IgE | Dizier et al., 1999; Nagarkatti et al., 2004 | ||
| 5q31 | IL-13 |
C-1055T; (C-1112T); A-1512C C1923T; G2525A; C2580A; C2749T; G427557A; +79Т>С; Arg110Gln |
Атопическая астма/ Общий IgE | Van der Pouw Kraan et al., 1999; Graveset al., 2000; Liu et al., 2000; Heinzmann et al., 2000; Eder et al., 2004 |
|
Β2-AR |
G-1023A; C-709A; G-654A; C-468G; C-406T; T-367C; T-47C; T-20C; G46A; C79G G252A; C491T; C523A; G-654A; G46A; Gly16Arg |
Лекарственный ответ (изучение гаплотипа) FEV1 Астма FEV1 Гормоно-зависимая астма |
Reihsaus et al., 1993; Drysdale et al., 2000; Summerhill et al., 2000 | |
| 5q31.1 | CSF2 | 117Thr | Астма | Hoffjan et al., 2004 |
| 5q31.1 | СD14 | -159C→T | Общий IgE | Baldini et al., 1999; Gao et al., 1999 |
| 5q35 | LTC4 synthase | -444C | Аспирин-зависимая астма | Senak et al., 2000 |
| 5q31 | SPINK5 | G1258A | Астма | Kabesch et al., 2004 |
| 6p | Il17F | Астма | Ramsey et al., 2005 | |
| 6p21 | HLA-II | Специфический IgE | Moffatt, 1996 | |
| 6p21.3 | TNF | G-308A | Астма | Sandford et al., 2004 |
| LT-α | Астма | Moffatt, Cookson, 1997 | ||
| 6р21-12 | PAF-acetyl-hydrolase |
Ile198Thr Ala379Val |
Атопическая астма/Общий IgE/ Специфический IgE |
Kruse et al., 2000 |
| 6p21 | HLA-G | Астма/ Бронхиальная гиперреактивность | Nicolae et al., 2005 | |
| 7р | Гаплотип-блок GPR154 (GPRA) | Астма | Melen et al., 2005 | |
| 8p23.1-p21.3 | NAT2 | 590G>A | Астма | Ляхович и др., 2000 |
| 9q33.1 | TLR4 | Asp299Gly | Астма | Fageras Bottcher et al., 2004 |
| 10p15 | GATA3 |
Фенотипы астмы |
Pykalainen et al., 2005 | |
| 11q12-13 | CC16 | A38G | Астма | Laing et al., 1998 |
| 11q12.1 | FcεR1-β |
237Gly Ile181Leu Leu181/Leu183 E237G |
Астма/ Атопия/ Бронхиальная гиперреактивность | Shirakawa et al., 1994, 1996; Hill et al., 1995, 1996; Hoffjan et al., 2004 |
| 11q13 | GSTP1 |
Ile105Val |
Атопическая астма/ IgE/ Кожные аллергопробы/FEV1/ Атопический дерматит |
Fryer et al., 2000; Cафронова и др. 2003; Tamer et al., 2004 |
| 12q13 | STAT6 | G2964A | Астма | Gao et al., 2000 |
| 12p13.31 |
C3 C3AR1 |
4896C/T 1526G/A |
Астма/ Общий IgE |
Hasegawa et al., 2004 |
| 12q24.2-q24.31 | NOS1 | 5266 C/T | Общий IgE | Holla et al., 2004 |
| 13q14.2 | CYSLTR2 |
601A>G; -1220A > C |
Астма | Pillai et al., 2004; Fukai et al., 2004 |
| 13q14 | PHF11 | Атопический дерматит | Jang et al., 2005 | |
| 14p11 |
TCR |
VA8.1(*)2 | Специфический IgE | Moffatt et al., 1997 |
| 14q32 | TLR2 | TLR2/-16934 | Атопия | Eder et al., 2004 |
Mast cell chymase |
MCC BstXI |
Экзема/ Общий IgE/ Атопический дерматит |
Mao et al., 1996, Tanaka et al., 1999 | |
| 16p12.1 | IL4R | Гаплотип C-3223T, Q551R, I50V | Атопическая астма | Hytonen et al., 2004 |
| 16р12-р11 | IL-4Rα |
Q576R; S503P; Ile50Val; Ser727Ala; Glu375Ala;Cys406Arg;Ser411Leu;Ser478Pro;Ser761Pro;Gln551Arg;T/C (+22446) |
Атопия/IgE/ Атопический дерматит/Атопическая астма (исследование гаплотипа) |
Hershey et al., 1997; Mitsuyasu et al., 1998; Kruse et al., 1999; Ober et al., 2000; Adjers et al., 2004 |
| 17q11-q12 |
RANTES |
G403A | Астма/ атопия/ атопический дерматит | Nickel et al., 2000, Fryer et al., 2000 |
| 17q11.2 | NOS2A | D346D | Астма | Hoffjan et al., 2004 |
| 19q13.1 | TGFB1 | -509T allele | Астма | Hoffjan et al., 2004 |
| 22q11.2 |
GSTT1 |
del |
Астма/ Атопия |
Fryer et al., 2000, Вавилин и др., 2002, Иващенко и др., Ляхович и др., 2000 |
| Xq |
IL-13Rα1 |
A1398G | Общий IgE | Heinzmann et al., 2000 |
| DAP3 |
Астма/ Общий IgE |
Hirota et al., 2004 | ||
| LTC(4) | A(-444)C | Астма | Kedda et al., 2004 |
В отношении астмы проведено 13 полногеномных исследований (в том числе исследование в различных расовых группах), в результате чего было подтверждено сцепление БА с регионами 5q23-31, 6p21-23, 12q14-24, 13q21-qter и 14q11-13, а также определена важность новых регионов астмы 2q33, 5p15, 11p15, 17p11, 19q13, 21q21 [Daniels et al., 1996; The Collaborative Stady on the Genetics of Asthma, 1997, 2004; Ober et al., 1998; Hizawa et al., 1998; Wjst et al., 1999; Yokouchi et al., 2000; Cookson et al., 2001; Koppelman et al., 2002]. Полученные данные еще раз свидетельствуют в пользу того, что в этиологии и патогенезе БА задействовано исключительное множество генов, каждый из которых в отдельности может вносить лишь относительно небольшой вклад в общую генетическую подверженность к заболеванию.
Изучение этиологии и патогенеза БА показало важную роль в формировании этого заболевания интерлейкинов (ИЛ), ответственных за индукцию и поддержание воспаления при данной патологии [Chung, Barnes, 1999]. Интересным фактом оказалось то, что гены цитокинов, играющих существенную роль в патогенезе БА расположены тандемно в одном кластере на хромосоме 5q31-33 [Arai et al., 1990]. На сегодняшний день показана связь БА и ее клинических проявлений со многими генами ИЛ и их рецепторов [Nanavaty et al., 2001] Так, сотрудниками лаборатории популяционной генетики НИИ медицинской генетики (г.Томск) совместно с кафедрой факультетской педиатрии с курсом детских болезней (заведующий – д.м.н., профессор Огородова Л.М.) лечебного факультета Сибирского государственного университета в рамках работы по изучению генетической компоненты подверженности к БА была показана ассоциация аллеля С-703 гена IL5 с этим заболеванием, характеризующимся бронхиальной гиперреактивностью [Фрейдин и др., 2000]. Кроме того, установлено, что генотип G/G 3’-UTR гена IL4 является фактором риска тяжелого течения заболевания, а гетерозиготный генотип G/С 3’-UTR этого же гена – протективным фактором, ассоциированным с легкой астмой [Огородова и др., 2002; Freidin et al., 2003]. Анализ вклада генотипической изменчивости по генам ИЛ и их рецепторов в фенотипическое варьирование количественных, патогенетически значимых для БА признаков показал, что гены ИЛ и их рецепторов по отдельности определяют 2-5% общей фенотипической дисперсии количественных показателей (1,63-5,54% у мужчин и 1,03-2,15% у женщин) [Фрейдин и др., 2003].
К настоящему моменту накапливаются результаты работ, посвященных анализу связи полиморфизма генов системы ферментов метаболизма ксенобиотиков с атопическими заболеваниями (табл. 3). Многолетние исследования, проводимые в НИИ молекулярной биологии и биофизики СО РАМН (г. Новосибирск) и в НИИ акушерства и гинекологии им. Д.О. Отта (г. Санкт-Петербург) показали связь полиморфизма генов системы ферментов метаболизма с формированием предрасположенности к БА и особенностей ее клинического фенотипа.
Малочисленность имеющихся данных о значимости генов системы метаболизма ксенобиотиков для БА, а также их противоречивый характер, свидетельствуют о чрезвычайной актуальности таких исследований, так как относительно генов метаболизма в ряде случаев, возможно точно установить факторы, обусловливающие их патологический эффект [Ляхович и др., 2000; Schwartz et al., 2004].
Таблица 3
Связь полиморфных вариантов генов ферментов метаболизма ксенобиотиков с бронхиальной астмой и ее клиническими проявлениями
| Ген (полиморфизм) | Ассоциация | Литературный источник |
| GSTT1 (+/del) | БА/ пищевая аллергия | Ляхович и др., 2000; Иващенко и др., 2001; Вавилин и др., 2002; Gilliand et al., 2002; Brasch-Andersen et al., 2004 |
| GSTM1(+/del) | ||
| CYP1A1(Ile462Val) |
БА/ пищевая аллергия/ эозинофилия |
Ляхович и др., 2000; Вавилин и др., 2002 |
| NAT2(S1, S2) | БА/ пищевая аллергия/ эозинофилия |
Luszawaka-Kutrzela, 1999; Gawronska-Szklarz et al., 2001; Вавилин и др., 2002 |
| GSTP1(313A>G) | БА/ положительные прик-тесты/ уровень IgE/ атопический дерматит/ гиперреактивность бронхов | Fryer et al., 2000; Сафронова и др., 2003; Tamer et al, 2004; Сarroll, 2005 |
|
CYP2E1 (-2964G/A) |
Эффективность лечения БА |
Obase et al., 2003 |
Обобщая вышеизложенное необходимо отметить, что полученные данные позволили значительно продвинуться в определении генов, полиморфизм которых, возможно играет существенную роль в развитии заболевания. Однако вследствие сложного клинического фенотипа БА, полигенной модели наследования и значительной роли воздействий внешней среды в развитии и прогрессировании этого заболевания, большее число генов подверженности к астме до сих пор остается до конца не идентифицированным и требует дальнейшего исследования.
Туберкулез (ТБ) – одно из самых распространенных инфекционных заболеваний, характеризующееся преимущественно хроническим течением различных клинических форм, своеобразием специфических иммунологических и морфологических проявлений. Проникновение в организм возбудителя ТБ является необходимым, но недостаточным условием для развития болезни, и в патогенезе ТБ взаимосвязаны взаимодействие инфекционного агента, факторы среды и особенности организма хозяина (пол, возраст, сопутствующие заболевания, общая реактивность организма и т.д.). ТБ отличается клиническим полиморфизмом, который определяет различные формы заболевания – от малых с бессимптомным течением до обширных деструктивных процессов в легких с выраженной клинической картиной, а также наличием туберкулезного процесса различной локализации в других органах [Хоменко, 1990]. По-видимому, причины таких различий обусловлены не только неблагоприятным сочетанием внешних факторов, но и особенностями организма, обусловленными его генотипом. Так, отмечено, что некоторые индивиды проявляют врожденную относительную резистентность к ТБ [Авербах, 1976]. Благодаря этому заболевает лишь малая часть населения, в то время как, по данным ВОЗ, инфицируется практически каждый третий житель планеты [Шайхаев, 1999].
По результатам популяционных исследований были показаны этнические различия в развитии ТБ [Рудко и др., 2004; Cervino et al., 2000; Bellamy et al., 1998]. Возможно, что этнические различия в предрасположенности к заболеванию обусловлены определенными традициями популяций, экономическими причинами и др. Кроме того, накоплены данные о высокой подверженности к ТБ популяций, происходящих с территорий свободных от этого заболевания: случаи заболевания были особенно высоки в популяциях, ранее не встречавшихся с этим заболеванием [Bellamy et al., 1998; Stead, 1992]. Эти данные подтверждают гипотезу, выдвинутую еще в 1949 г. Haldane о том, что инфекционные заболевания были главной силой естественного отбора, а резистентность к туберкулезной инфекции формировалась в процессе симбиотных отношений макро- и микроорганизмов [Земскова, 1984].
На сегодняшний день показана роль в подверженности к ТБ для многих генов, в том числе HLA-cистемы, NRAMP1, IFN-γ и его рецептора и др. Одним из главных генов-кандидатов туберкулеза является NRAMP1 (от англ. Natural-Resistance-Associated Macrophage Protein 1 gene – ген макрофагального белка, ассоциированного с естественной резистентностью 1) [Bellamy et al., 1998]. Белковый продукт этого гена Nramp1 участвует в процессах активации макрофагов, являясь ключевым звеном в механизме транспорта нитритов из внутриклеточных компартментов в более кислую среду фаголизосомы, где он способен вступать в химическую реакцию с образованием NO. Белок входит в семейство функционально связанных мембранных белков (к этому семейству относят также Nramp2), ответственных за транспорт двухвалентных катионов, таких как Fe2+, Mn2+, Zn2+, Cu2+ [Canonne-Hergaux et al., 1998]. Поэтому, нарушение работы системы, обеспечивающей транспорт важных веществ через мембрану, приводит к дисбалансу между выведением и поступлением веществ в клетки, что может способствовать изменению внутриклеточной концентрации ионов, вызывающей гибель клеток. Предполагаемый механизм антибактериальной функции Nramp1 лежит в создании неблагоприятной для бактерии окружающей среды внутри фагосомы [Пузырев и др., 2002]. Следовательно, дефекты продукции или функции Nramp1 могут приводить к нарушению его транспортной роли и, как следствие, к повышению чувствительности к внутриклеточным патогенам, таким как микобактерии.
Многочисленные работы на экспериментальных животных показали, что NRAMP1 играет важную роль в чувствительности к микобактериям и некоторым другим возбудителям инфекций у мышей, и вероятно, что его человеческий гомолог связан с подобными инфекциями у людей [North, Medina, 1998]. Для определения функции гена NRAMP1 в развитии ТБ были проведены исследования в различных популяциях: у западных африканцев в Гамбии, местного населения в Корее и Японии [Bellamy et al., 1998; Ryu et al., 2000; Gao et al., 2000]. В результате было обнаружено, что изменчивость данного гена связана с различиями в восприимчивости к ТБ. Связь полиморфных маркеров гена NRAMP1 в дальнейшем была подтверждена на семейном материале у больных ТБ родственных между собой индивидов, проживающих на территории Гвинеи-Конакри [Cervino et al., 2000]. Результаты клинико-генетических исследований и изучение ассоциаций ряда маркеров с заболеванием ТБ сформировали единое мнение, что восприимчивость к данной болезни находится под полигенным контролем, а отдельный вклад гена NRAMP1 – лишь небольшая доля в общей подверженности к инфекционному заболеванию [North, Medina, 1998].
Другой генетической системой, задействованной в возникновении и патогенезе ТБ, считается комплекс HLA (от анг. Human Leukocyte Antigens). Комплекс HLA, как и его аналоги у животных, называют главным комплексом гистосовместимости, поскольку первой из обнаруженных функций этого комплекса был контроль над трансплантационным иммунитетом. Следует отметить, что комплекс генов HLA является чрезвычайно полиморфной системой. Первой работой в области исследования HLA системы при ТБ, где были получены положительные результаты, была работа R. Selby и соавт. (1978). Затем исследования были продолжены на популяционном и семейном материале, в ходе которых были получены противоречивые результаты. Ассоциации, показанные в одних популяциях, не находили своего подтверждения у других. В 1979 г. Al-Arif с соавт. показали значимое повышение встречаемости у больных ТБ легких антигена В15 в популяции американских негров [Al-Arif et al., 1979]. Позднее Jiang с соавт. обнаружили высокую частоту встречаемости антигена HLA-В27 среди заболевших ТБ китайцев [Jiang et al., 1983].
Отечественными исследователями также была проведена большая работа по изучению значимости HLA системы при заболеваемости ТБ в разных этнических группах, проживающих на территории России. В ходе работы была установлена повышенная частота встречаемости антигенов локуса HLA-B12,-С в узбекской и туркменской популяциях. У русских с ТБ легких в локусе HLA-В антигены В5, В14 встречались значимо чаще, но особенно интересным показался тот факт, что во всех трех популяциях показана ассоциация HLA-C локуса с заболеванием [Литвинов и др., 1983, 1986; Хоменко и др., 1985]. Следовательно, во всех изучаемых популяциях установлена взаимосвязь между некоторыми генетическими маркерами системы HLA и восприимчивостью к туберкулезу, причем в разных популяциях – с разными антигенами. В большинстве исследуемых популяций определены ассоциации заболевания ТБ с одним и тем же антигеном локуса DR (DR2), и учитывая, что гены комплекса HLA-DR отвечают за иммунный ответ, предполагается, что данный локус оказывает влияние на восприимчивость к ТБ, регулируя силу иммунного ответа на микобактериальные антигены [Поспелов и др., 1987; Хоменко, 1990]. В целом, гены, составляющие комплекс HLA, являются важными факторами патогенеза данного инфекционного заболевания. Об этом свидетельствует целый ряд многократно подтвержденных фактов: ассоциация определенных генов HLA (преимущественно DR и B-локусов) с заболеванием в большинстве обследованных популяций, сцепление гаплотипов HLA в семьях с пораженными родителями и детьми, ассоциации со специфичными антигенами у больных с хроническим, плохо поддающимся лечению процессом.
К настоящему моменту роль в подверженности к ТБ для генов рецептора к витамину D, γ-интерферона и его рецептора, фактора некроза опухолей, интерлейкинов и др. не вызывает сомнения [Bornman et al., 2004]. Интерес к системе генов метаболизма ксенобиотиков в отношении ТБ обусловлен несколькими причинами. Во-первых, данные, что при воспалении и инфекции происходит изменение уровня активности цитохромов Р450, предполагают задействование системы метаболизма в защите организма от последствий развертывания воспалительных реакций при заболевании [Prandota, 2002; Сибиряк, 2003; Бикмаева и др., 2004]. Во-вторых, знания об участии ферментов системы метаболизма в биотрансформации лекарственных препаратов, позволяют найти и избежать причины, определяющие нежелательные проявления терапевтического действия лекарств. В этом контексте определение генетической компоненты подверженности к проявлению многообразных побочных реакций при применении антимикобактериальных препаратов при ТБ имеет очень важное значение для достижения успехов в терапии заболевания [Dickinson et al., 1981; Roy et al., 2001; Huang et al., 2002, 2003].
Сложность патогенеза, а так же различия в клиническом проявлении ТБ предполагают, что число генов-кандидатов заболевания достаточно велико, при этом вклад каждого из них в суммарную подверженность различен [Hill, 1998]. И поэтому изучение полиморфизма известных генов-кандидатов, а также поиск новых генов, белковые продукты которых в той или иной степени вовлечены в патогенетические механизмы заболевания, представляется одной из приоритетных задач.
1.3. Полиморфизм генов ферментов биотрансформации ксенобиотиков и патология
Известно, что многие бронхолегочные патологии в различной степени связаны с развитием окислительного стресса. Эпителий легкого, насыщенного кислородом внешней среды, чрезвычайно восприимчив для токсического действия радикалов экзогенного и эндогенного происхождения. Высокая частота заболеваний бронхолегочной системы (астма, эмфизема, пневмония и др.) находится в прямо пропорциональной зависимости от уровня загрязнения окружающей среды сильными окислителями (NO, NO2, CO, O3, альдегиды), пылевыми частицами в совокупности с воздействием экстремальных климатических условий [Гусев, Даниловская, 1987; Mutmansky, 1990; Тиунов и др., 1991]. Состояние окислительного стресса и разрушающее воздействие свободнорадикального окисления имеет значение не только в возникновении заболевания, а также может являться важнейшей причиной дальнейшей хронизации патологического процесса в легочной ткани [Меньщикова, Зенков, 1991].
Известно, что источниками активированных кислородных метаболитов могут быть как внешние факторы (альдегиды, озон, окислы азота, сигаретный дым, анаэробные бактерии), так и эндогенные, задействованные во внутриклеточных метаболических процессах (альвеолярные макрофаги, гранулоциты, внутриклеточные органеллы). Воздействие атмосферных прооксидантных поллютантов, таких как озон, окислы азота, составляющих табачного и автомобильного дыма на дыхательные пути приводит к индуцированию окислительных процессов, как на поверхности бронхоальвеолярного секрета, так и непосредственно в эпителии легкого [Wright et al., 1994]. Присутствие разнонаправленных повреждающих воздействий оксидативного стресса говорит о важности для организма поддержания баланса системы активированных кислородных метаболитов в легких.
Эффективной защитой от различных токсикантов внешней среды, поступающих с вдыхаемым воздухом, служит система биотрансформации ксенобиотиков при согласованном функционировании защитных механизмов. Глутатион S-трансферазы семейство ферментов, участвующих в метаболизме большого числа электрофильных ксенобиотиков через конъюгацию с глутатионом, а также в метаболизме ряда эндогенных субстратов (гормонов, липидов, простагландинов, лейкотриенов). Таким образом, метаболизм ксенобиотиков через глутатионопосредованную детоксикацию играет важную роль в обеспечении устойчивости клеток к перекисному окислению жиров, свободным радикалам, алкилированию белков, в формировании резистентности к лекарственным препаратам и предотвращении поломок ДНК.
В результате однонуклеотидной замены аденина (А) на гуанин (G) в гене GSTP1, приводящей к замене аминокислот изолейцина (Ile105) на валин (Val105), происходит изменение ферментативной активности, обусловливающее повышение уровня гидрофобных аддуктов в тканях легких и полициклических ароматических углеводородов-ДНК аддуктов в лимфоцитах крови. Было выявлено, что замена изолейцина на валин в 105 положении расположенная в субстрат-связывающем Н участке фермента, приводит к различным изменениям кинетических параметров фермента [Katoh et al., 1999]. Показано, что при мутации Val105 в 7 раз увеличивается каталитическая активность фермента по отношению к полициклическим ароматическим соединениям, но в 3 раза снижается активность по отношению к 1-хлор-2,4-динитробензену [Ishii et al., 1999]. Отмечено, что индивидуумы с аллелем Val105 имеют повышенный риск развития РЛ [Баранов и др., 2000].
К настоящему времени накоплено достаточно сведений об ассоциации «нулевого» генотипа гена GSTM1 с риском развития эмфиземы легких и хроническим бронхитом у курильщиков [Афанасьева, Спицин, 1990], кроме того, показана повышенная частота «нулевого» генотипа, помимо GSTM1, и для гена GSTT1 у больных БА [Баранов и др., 2000]. Микросомальная эпоксигидролаза (EPHX1) осуществляет метаболизм продуктов табачного дыма, и поэтому играет важное значение в защите легких от высокоактивных производных эпоксида, образующихся при курении и приводящих к повреждению легочной ткани курильщиков. Показано, что с аллелем S гена EPHX1, обеспечивающим пониженную активность соответствующего фермента, ассоциированы заболевания органов дыхания, такие как эмфизема легких, хронический обструктивный бронхит, муковисцидоз, хронические респираторные заболевания [Баранов и др., 2000; Lomas, Silverman, 2001; Matsushita et al., 2002; Sandford, Silverman, 2002].
Многочисленные исследования полиморфных вариантов генов системы метаболизма ксенобиотиков показали связь с различными заболеваниями, включая сердечно-сосудистую патологию, атопические заболевания, хронические неспецифические заболевания легких и др. Но, прежде всего, пристальное внимание исследователей к индивидуальным особенностям функционирования системы биотрансформации отмечено при онкологических заболеваниях. Это понятно, так как уже доказано влияние большинства химических агентов, с которыми человеку приходится сталкиваться как в быту, так и на производстве, на процессы канцерогенеза.
Исследование полиморфизма 313 A>G гена GSTP1 у японцев с различными онкологическими патологиями (рак ротовой полости, легких, желудка, колоректальным и урогенитальным видами рака) показало ассоциацию только у некурящих индивидуумов с раком ротовой полости (РРП), для остальных видов рака различий не было выявлено [Katoh et al., 1999]. В другой работе была изучена роль полиморфного варианта C341T гена GSTP1 как важного фактора, обусловливающего развитие РРП, где был показан повышенный риск развития данной патологии, как для европеоидов, так и для афроамериканцев. Интересен тот факт, что более высокий риск развития заболевания наблюдался у пациентов с малым потреблением табака (< или = 20 пачка/год) по сравнению с группой интенсивных курильщиков (>20 пачка/год) [Park et al., 2000].
Генетическая предрасположенность – одна из важных гипотез, объясняющих, почему лишь у малого числа курильщиков развивается рак легкого (РЛ). Полиморфизмы, участвующие в метаболизме канцерогенов изучаются как факторы риска для рака легкого. Полиморфизм в гене GSTM1, также как и в гене GSTT1, обусловлен протяженной делецией, в результате которой происходит полное отсутствие ферментативной активности. В ряде работ показано, что функциональную значимость в развитии онкопатологии может иметь не один конкретный рисковый генотип, а их специфическая комбинация. Было отмечено, что повышенный риск РЛ у курильщиков европеоидной расы имеют индивиды с комбинацией генотипов GSTM1-нуль, GSTP1 AG+GG и GSTM3 AA (n=322) [Jourenkowa-Mironowa et al., 1998]. Другое исследование также выявило связь развития РЛ у индивидов с «нулевым» генотипом гена GSTM1 в присутствии р53 Pro аллеля [Miller et al., 2002]. Предположительно, что потенциальное взаимодействие между GSTP1 и GSTM1 генами в японской популяции у мужчин-курильщиков (n=542) в возрасте 50-69 лет приводит к повышенному риску РЛ при комбинации одного из вариантов аллелей GSTP1 и нулевого генотипа гена GSTM1 [Kihara, Noda, 1999]. Выявлен повышенный риск РЛ среди курильщиков в популяционных выборках Средиземноморья, 93% которых составили мужчины с комбинацией «нулевого» генотипа гена GSTM1 и р53 Pro/Pro+Arg/Pro генотипов [To-Figueras et al., 1996].
Предположительное влияние изотиоцианатов – компонентов, обладающих антиканцерогенными свойствами и содержащихся в крестоцветных овощах, в снижении регуляции уровня ферментов биотрансформации семейства цитохромов Р450 и индукции ферментов второй фазы детоксикации, легло в основу исследования GSTM1 и GSTT1 генотипов у больных с впервые выявленным РЛ. В ходе исследования было показано модифицирующее влияние употребления в пищу изотиоцианатов у курильщиков гомозиготных по «нулевым» генотипам GST на развитие рака легкого [Spitx et al., 2000].
Миелодиспластический синдром (МДС) – клональное пролиферативное нарушение костного мозга, часто прогрессирующее в острую миелоидную лейкемию (ОМЛ), а заболеваемость и смертность от МДС одинакова и составляет период менее года. Воздействие различных по качественному и количественному составу химических веществ, с которыми приходится сталкиваться индивидуумам по роду профессиональной деятельности, на процессы канцерогенеза может увеличивать вероятность заболеваемости МДС. Повышенный риск МДС отмечен у лиц, проходивших курс химиотерапии при лечении других опухолей. Вследствие индивидуальных различий в метаболизме ксенобиотиков (в том числе и канцерогенов) возможно увеличение риска раковых заболеваний при сниженной функции метаболизирующих ферментов. Конъюгация электрофильных компонентов с глутатионом, опосредованная глутатион S-трансферазами, является важным этапом метаболизма канцерогенов. Известно, что частота «нулевого» генотипа гена GSTM1 среди европеоидов составляет порядка 50% и данный генотип ассоциирован с развитием РЛ, индуцированного курением, а также раком мочевого пузыря. О взаимосвязи «нулевого» генотипа гена GSTT1 с различными раковыми заболеваниями известно намного меньше, но ясно, что у индивидуумов с «нулевым» генотипом снижена способность к метаболизму некоторых канцерогенов, включая 1,3-бутадиен, метилбромид, оксид этилена. Был установлен четырехкратный относительный риск МДС для носителей «нулевого» генотипа GSTT1 [Chen et al., 1996].
Предполагается, что нулевой генотип гена GSTT1, ассоциированный с канцероген-индуцированными хромосомными изменениями в лимфоцитах, может увеличивать риск подверженности к миелодисплазии. При анализе данных, полученных в ходе исследования пациентов с острой миелоидной лейкемией (ОМЛ), была показана повышенная частота делеции в гене GSTT1 среди больных (60%), что практически в три раза выше, чем в контрольной группе (17%), кроме того, индивиды с делецией по генам GSTT1 и GSTM1 имели несколько больший риск ОМЛ. Интересно, что у лиц со вторичной ОМЛ делеция GSTT1 встречается на 20% чаще по сравнению со случаями de novo, что характерно и для индивидов с «нулевым» генотипом гена GSTM1 [Cramp et al., 2000].
Возможно, что эффекты определенных генотипов генов ферментов биотрансформации различны в развитии рака и других заболеваний. В связи с этим предложено, что продукты, образующиеся в ходе метаболизма ряда ксенобиотиков с участием глутатионовых S-трансфераз, способствуют атерогенезу и нестабильности тромбоцитов. В дальнейшем было показано, что «нулевой» генотип гена GSTM1 играет протективную роль в развитии инфаркта миокарда, причем эффект более выражен у курильщиков [Wilson et al., 2000]. Вероятно, что наличие «нулевого» генотипа по генам GST способствует повышенной регуляции других ферментов, более эффективно участвующих в метаболизме атерогенных субстратов с учетом одного из важных качеств системы биотрансформации, а именно широкой субстратной специфичности. По этому поводу имеются данные о скоординированной экспрессии GSTM1 и GSTM3 в легочной ткани человека [Anttila et al., 1995], а также о более высокой активности CYP1A2 у индивидуумов с нулевым генотипом GSTM1 [MacLeod et al., 1997].
Глутатионопосредованная детоксификация принимает непосредственное участие в защите организма от оксидативного стресса, что оправдывает изучение полиморфизма генов глутатион S-трансфераз в патогенезе различных патологических состояний, в том числе и при эндометриозе. Так, у больных эндометриозом женщин Башкортостана отмечаются различия по частотам как отдельных генотипов полиморфных локусов GSTM1 и GSTP1, так и по распределению частот их сочетаний. Кроме того отмечено, что более выраженный эффект от гормонального лечения наблюдался у лиц, имеющих «нулевой» генотип гена GSTM1 в сочетании с мутацией по гену GSTP1 [Шарафисламова и др., 2003].
N-ацетилтрансфераза-2 (NAT2) и микросомальная эпоксигидролаза (EPHX1) полиморфные гены ферментов, метаболизирующих различные канцерогены табачного дыма. Табачный дым содержит >4000 компонентов, включая около 50 канцерогенов, являющихся субстратами семейства ферментов биотрансформации. Для понимания роли этих двух полиморфизмов во взаимодействии «ген-окружающая среда» в развитии РЛ было проведено исследование, в ходе которого было обнаружено, что риск развития заболевания значительно повышается с увеличением значения «пачка-лет» у курильщиков. Результаты данного исследования еще раз показали очевидность того, что изучение взаимоотношения генетического полиморфизма с факторами окружающей среды в формировании повышенного риска онкологической патологии, имеет состоятельность только тогда, когда внешнесредовой фактор является неотъемлемой составляющей частью патогенетического механизма (например, курение), и он обязательно включается в анализ [Zhou et al., 2002].
Ген CYP17 кодирует фермент цитохром Р450С17a, который выполняет две различные функции в биосинтезе стероидов, что обусловливает его изучение как гена кандидата восприимчивости к эндокринзависимым опухолям. Тем не менее, были получены довольно противоречивые результаты при исследовании пациентов с раком яичников и полиморфизма Т-С в промоторном регионе CYP17 гена в различных популяциях [Spurdle et al., 2000]. Что еще раз подтверждает популяционную вариабельность полиморфизма генов биотрансформации, и, следовательно, различный вклад генов системы метаболизма у разных индивидуумов в процессы онкогенеза.
Ген CYP1A1 человека был клонирован и секвенирован в 1986 году и локализован на хромосоме 15. Полиморфизм в 3’-некодирующем регионе гена, обусловленный заменой цитозина на тимидин, узнаваемый MspI эндонуклеазой рестрикции был впервые идентифицирован у японцев. Различные исследования показали ассоциацию данного полиморфизма и риском развития РЛ в европеоидной популяции [Shields et al. 1993; Alexandrie et al., 1994; Sugimura et al., 1994]. Эти результаты сходны с данными, полученными в аналогичной работе по изучению злокачественного новообразования в легких для японской популяции [Xu et al., 1996].
Употребление алкоголя в больших дозах рассматривается как один из факторов, способствующих развитию различных заболеваний печени. Так, отмечен высокий риск развития гепатоцеллюлярной карциномы (ГЦК) у японцев, злоупотребляющих алкоголем. Причем, наблюдалась повышенная частота С2 аллеля гена CYP2E1, связанного с высокой транскрипционной и ферментативной активностью, что также позволяет говорить о повышенном риске к ГЦК у индивидуумов, злоупотребляющих алкоголем [Munaka et al., 2003].
Развитие алкогольного поражения печени (АПП) на фоне алкогольной интоксикации всего организма является следствием несостоятельности ферментативной системы биотрансформации ксенобиотиков, участвующей в метаболизме этанола. Метаболизм этанола происходит в печени, где метаболизируется порядка 98% попавшего в организм алкоголя по НАДФ-зависимому пути с помощью алкогольдегидрогеназы и ацетилдегидрогеназы, локализованных преимущественно в цитоплазме клеток печени. Существует и другой путь окисления этанола с помощью микросомальной этанолокисляющей системы при участии ферментов семейства цитохрома Р450. Шангареева З.А. и др. показали, что для пациентов, страдающих АПП характерно повышение частот мутантных аллелей генов CYP2E1, CYP1A1, mEPHX, GSTT1 и GSTM1, приводящее к увеличению рисковой значимости гетерозиготных генотипов генов CYP2E1 (OR=7,37), CYP1A1 (OR=2,87), mEPHX (OR=2,45) [Шангареева и др., 2003].
Начало или обострение псориаза, обусловленного Т-клеточным механизмом заболевания кожи с аутоиммуным характером заболевания, зачастую вызывается применением b-блокаторов и противомалярийных лекарственных препаратов. Предполагается, что метаболическая эффективность, обусловленная различными вариантами аллелей генов системы ферментов биотрансформации, может привести к накоплению ксенобиотиков или их реактивных метаболитов в органах-мишенях, а в дальнейшем неоантигены или неизвестные пептиды могут вызвать агрессивную реакцию со стороны Т-клеток. В этом контексте, было проведено исследование полиморфизма гена CYP2C19 у пациентов с псориазом. В ходе исследования было показано, что для гетерозиготных носителей по гену CYP2C19 (*1А и *2А) характерно более позднее развитие псориаза, в то время как эти же генотипы показали протективную роль для развития артрита, связанного с псориазом [Richter-Hintz et al., 2003].
Цитохромы P450 ответственны приблизительно за 75% метаболизма лекарственных препаратов и различных химических агентов. Человек имеет 59 активных генов, и 6 из них кодируют важные для лекарственного метаболизма ферменты. Приблизительно 40% цитохром P450-зависимого лекарственного метаболизма катализируются полиморфными ферментами, и такие «лекарство→P450» взаимодействия часто рассматриваются в отношении побочных действий лекарственных препаратов [Ingelman-Sundberg, 2004].
Экспрессия цитохрома P450 и связанная с ней биотрансформация изменяется при различных инфекционных заболеваниях. Следовательно, при развитии воспаления и инфекции в организме нарушена способность метаболизма печени и других органов, контролирующих действие лекарств. Содержание цитохрома Р450 и монооксигеназные активности в тканях этих органов снижаются при развитии бактериальных и вирусных инфекций, иммунизации различными антигенами, в условиях фармакологической иммуностимуляции, что опосредовано цитокинами. Депримирующее воздействие на цитохром Р450-зависимые монооксигеназы обнаружено у IFNa, IFNb, IFNg, IL-1 и TNF, IL-6, IL-11, IL-2. Цитокины угнетают транскрипцию генов и накопление мРНК различных изоформ цитохрома Р450 в клетках, возможно и посттранскрипционное угнетение синтеза белка некоторых изоформ [Renton, 2004]. Благодаря цитокиновой модуляции процессов монооксигенирования на цитохроме Р450, реализуется адаптация механизмов биотрансформации низкомолекулярных ксенобиотиков в условиях активации иммунной системы. Это обеспечивает, с одной стороны, защиту от последствий возможной неконтролируемой активации потенциально опасной для организма ферментной системы и снижение риска нарушения оксидантного равновесия, с другой - сохранение оптимального уровня и неогенез необходимых для адекватного "ответа" организма низкомолекулярных липофильных сигнальных молекул. Угнетение фармакометаболизирующей функции печени и изменение фармакодинамики и токсичности лекарственных препаратов необходимо учитывать при проведении терапии препаратами рекомбинантных цитокинов. В этом случае необходимо отметить, что ингибирование метаболизма различными препаратами также как влияние на концентрацию и/или число различных цитокинов в воспаленных тканях, может вызывать положительные эффекты у пациентов с различными заболеваниями, что позволит говорить о новых терапевтических возможностях лекарственных средств [Сибиряк, 2003].
Разнообразие элементов многокомпонентной и многоэтапной системы метаболизма имеет важное значение для фармакогенетики в плане разработки индивидуального подхода к лечению пациента. C помощью предупреждения индивидуального ответа на лекарственный препарат становится возможным повышение эффективности лечения и устранения нежелательных эффектов от медикаментозной терапии [Баранов и др., 2000].
Заключая обзор в целом, необходимо отметить, что изучение естественной изменчивости генов ферментов биотрансформации ксенобиотиков у больных и членов их семей, а также установление их вклада в патогенез распространенных заболеваний, таких как БА и ТБ, стало задачей настоящего исследования. А понимание основных механизмов, участвующих в патогенезе заболеваний, поможет понять не только причины возникновения болезней, но и научиться бороться с ними.
ГЛАВА 2. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
2.1. Характеристика обследованных групп населения
В рамках проведенного исследования был проанализирован полиморфизм генов биотрансформации ксенобиотиков и уточнена их функциональная значимость в отношении БА и ТБ легких у русских жителей г. Томска.
Выборки были сформированы для данного исследования на основе ДНК-банка лаборатории популяционной генетики НИИ медицинской генетики ТНЦ СО РАМН, созданного сотрудниками этой лаборатории.
Контрольная группа.
В качестве контрольной группы использовалась выборка, принадлежащая в настоящее время банку ДНК лаборатории популяционной генетики НИИ медицинской генетики ТНЦ СО РАМН. Выборку в количестве 140 человек (средний возраст ±S.D. 64,3±18,0) частично составили индивиды не родственные между собой и не имеющие по результатам клинического обследования бронхолегочной патологии. Среди индивидов контрольной группы было 80 женщин (средний возраст ±S.D. 65,8±17,8) и 60 мужчин (средний возраст ±S.D. 63,5±18,1).
2.1.1. Характеристика группы больных туберкулезом
Материал для исследования больных ТБ был собран на базе Областной Томской Клинической туберкулезной больницы, Детского легочно-туберкулезного отделения Железнодорожной больницы, Областной детской туберкулезной больницы, а также Областном противотуберкулезном диспансере, туберкулезного отделения Областной психиатрической больницы. Сбор материала и клиническое обследование больных осуществлялось при участии сотрудников кафедры фтизиатрии Сибирского государственного медицинского университета (заведующий – член–корр. РАМН, профессор Стрелис А.К.). Основным критерием отбора в группу пациентов были два условия – отсутствие родственных связей между индивидами и этническая принадлежность.
Общая выборка больных туберкулезом легких.
Выборку составили 304 индивида, средний возраст ±S.D. которых был 30,6±15,4, из них 99 женщин, средний возраст ±S.D. которых составил 26,3±14,6 года и 205 мужчин средний возраст ±S.D. – 32,8±15,4 лет. Диагноз туберкулеза легких устанавливался на основании данных микроскопии мокроты с обязательным рентгенологическим исследованием легких для определения формы заболевания и распространенности специфического процесса (общепринятые методы). Противотуберкулезную терапию больные получали по 1-ой категории, согласно рекомендациям ВОЗ (табл. 4).
Таблица 4
Режимы лечения больных с распространенным деструктивным туберкулезом легких по протоколам ВОЗ
|
Масса тела до начала лечения, кг |
Начальная фаза: 2 месяца |
Фаза продолжения |
|||
| 4 месяца | |||||
|
Иониазид+ Рифампицин, таблетка 100мг+150мг 150мг+300мг |
Пиразинамид, таблетка 500 мг |
Этамбутол, таблетка 400мг |
Стрептомицин, порошок для инъекций, 1г основания во флаконе |
Иониазид+ Рифампицин, таблетка 100мг+150мг 150мг+300мг |
|
| 33-50 |
3 табл. (100мг+150мг) ежедневно |
3 табл. ежедневно |
2 табл. ежедневно |
750 мг ежедневно |
3 таблетки ежедневно |
|
50 и более |
2 таблетки (150 мг+ 300мг) ежедневно |
4 табл. ежедневно |
3 табл. ежедневно |
1 г ежедневно |
4 таблетки ежедневно |
Семейная выборка больных туберкулезом легких.
Семейная выборка была зарегистрирована по пробандам – больным туберкулезом, находившихся на лечении в противотуберкулезных учреждениях г. Томска в период с 2000 по 2004 г.. Всего было обследовано 42 семьи (109 человек), в том числе 25, зарегистрированных по пробандам – детям в возрасте от 1 года до 15 лет (средний возраст ±S.D. составил 7,7±3,9). Семнадцать семей было выбрано по взрослым пробандам в возрасте от 17 до 48 лет (29,4±12,3 лет). В составе «пробанд/пробанды-мать-отец» исследовано 19 семей, в неполном составе, когда отсутствовал материал одного из родителей – 16 семей.
Группу пробандов – детей составили 10 мальчиков (7,2±2,9 лет) и 15 девочек (7,9±4,5 лет). Средний возраст пробандов – детей разного пола достоверно не различался (р>0,05). Среди взрослых пробандов было 7 женщин (19,8±7,9 лет) и 10 мужчин (23,9±9,1 лет). Всем пробандам был поставлен диагноз туберкулеза.
2.1.2. Характеристика группы больных атопической бронхиальной астмой
Исследованная семейная выборка была зарегистрирована по пробандам больным БА, находившихся под наблюдением в клинико-профилактических учреждениях г. Томска в 1997-2000 г.. Клиническое обследование и диагностику провели сотрудники кафедры факультетской педиатрии с курсом детских болезней (заведующий д.м.н., профессор Огородова Л.М.) Сибирского государственного медицинского университета, областного детского центра клинической иммунологии и аллергологии (Областная детская больница, г. Томск, главный врач – Сальников В.А.).
Семейная выборка больных БА.
Всего обследовано 76 семей (213 человек), 61 семья из которых была набрана в составе трех человек «пробанд-мать-отец» и 15 семей – в составе двух человек, т.е. «пробанд-мать/или отец». В исследуемой выборке были 72 индивида, зарегистрированные по пробандам-детям в возрасте от 1,7 до 15 лет (средний возраст ±S.D.составил 8,5±3,5). Восемь семей было выбрано по взрослым пробандам в возрасте от 24 до 42,5 лет (34,4±7,2 лет).
Основную часть пробандов-детей составили мальчики (n=47), девочек примерно в два раза меньше (n=25). Средний возраст пробандов-детей разного пола достоверно не различался (8,4±3,5 лет у мальчиков и 8,6±3,5 лет у девочек; р>0,05). Среди взрослых пробандов было семь женщин (34,4±7,2 лет). Всем пробандам был поставлен диагноз «атопическая бронхиальная астма».
Выборка больных БА.
Выборку больных БА составили 134 индивида (21,9±18,8 лет), из них 59 - женщины в возрасте от 2 до 79 лет (24,7±18,5 лет) и 75 – мужчины в возрасте от 1,7 до 68 лет (19,7±18,9 лет). Среди больных БА 78 индивидов – дети в возрасте от 1,7 до 15 лет (8,8±3,7 лет), из которых 28 девочек в возрасте от 2 до 15 лет (8,8±3,9 лет) и 50 мальчиков в возрасте от 1,7 до 14,8 лет (8,6±3,5 лет). Кроме того, среди индивидов общей выборки больных БА было 56 взрослых (40,3±15,6 лет), из которых 31 женщина в возрасте от 19 до 79 лет (39,0±13,9 лет) и 25 мужчин от 19 до 68 лет (41,9±17,5 лет). Между группами мальчиков и девочек, а также мужчин и женщин не показано различий по возрастному критерию (р>0,05). Всем пациентам был выставлен диагноз «атопическая бронхиальная астма».
2.2. Характеристика методов исследования
2.2.1. Клинико-лабораторные методы
Пробанды, а также их родственники первой степени родства, согласившиеся на проведение исследования, были обследованы для верификации диагноза БА и симптомов атопии. Обследование включало сбор семейного анамнеза и многочисленные клинические тесты: в данной работе были использованы результаты только спирометрических, аллергологических и иммунологических анализов.
Диагноз «бронхиальная астма» верифицировали на основании критериев ВОЗ: наличие характерного для заболевания анамнеза, типичных клинических симптомов астмы, атопии (атопический анамнез, положительные скарификационные аллергопробы (САП), уровень общего сывороточного IgE более 100 МЕ/мл) [Бронхиальная астма. Глобальная стратегия, 1996]. В случае невозможности доказать наличие атопии, выставляли диагноз неатопической БА. Степень тяжести заболевания устанавливали согласно критериям проекта GINA (2002 г.) и Национальной программы лечения и профилактики БА у детей (1997 г.).
Аллергологическое обследование включало сбор аллергоанамнеза и проведение САП на пищевые, ингаляционные, эпидермальные, растительные и грибковые аллергены с использованием стандартных наборов согласно рекомендациям производителей («Биомед», Москва; «Immuno Tek», Испания).
Измерение уровня общих сывороточных антител класса E проводили с помощью твердофазного иммуноферментного анализа с использованием стандартных наборов согласно рекомендациям производителей («Протеиновый контур», Санкт-Петербург; «Veda Lab», Франция). Уровень общего сывороточного IgE пересчитывали на международные единицы на миллилитр (МЕ/мл; 1 МЕ =2,42 нг/мл).
Исследование функции внешнего дыхания (ФВД) осуществляли по стандартной методике (анализ кривой «поток-объем» и показателей спирометрии) на установке «Master Lab Pro» («Эрих Йегер», Германия) [Quanjer et al. 1993].
Для определения степени реактивности бронхов проводили провокационный тест с метахолином. Диапазон концентраций растворов метахолина составили 0,25-32 или 64 мг/мл. Результаты выражали как концентрация метахолина, вызывающая не менее чем 20% падение объема форсированного выдоха (РС20), вычисленная методом линейной интерполяции по общепринятой формуле [Sterk et al., 1993]. Диагностически значимой в отношении БА считали РС20 не менее 20 мг/мл – это состояние рассматривали как наличие бронхиальной гиперреактивности (BHR).
В отношении больных ТБ был проведен полный клинико – эпидемиологический анализ с учетом возраста начала заболевания, социального статуса, вредных привычек (курение, злоупотребление алкоголем, употребление наркотиков), сопутствующей патологией, наличия контакта с туберкулезным больным, а также данные о ТБ в роду. Анализу подвергались выраженность клинических проявлений (жалобы, объективный статус больного), результаты лабораторных и инструментальных методов исследования (микроскопия и посев мокроты на МБТ, чувствительность к противотуберкулезным препаратам, рентгенологическое исследование легких) на момент начала заболевания, а также через 2 месяца лечения.
Определение количества эритроцитов, концентрации гемоглобина, общего числа лейкоцитов и их отдельных морфологических форм, величину СОЭ, уровень печеночных проб (билирубина, аланинаминотрансферазы и аспартатаминтрансферазы) исследовали общепринятыми методами [Меньшиков, 1987].
2.2.2. Молекулярно-генетические методы
В ходе выполнения работы было исследовано 6 полиморфных вариантов генов цитохромов Р450 (CYP2C19, CYP2E1) и глутатионовых S-трансфераз (GSTT1, GSTM1 и GSTP1) (табл. 4).
Для генотипирования индивидов по указанным полиморфизмам использовали образцы тотальной ДНК из банка НИИ медицинской генетики (семейная выборка больных БА и контрольная выборка) и ДНК, выделенную из цельной венозной крови (семейная выборка больных ТБ) по стандартной неэнзиматической методике [Маниатис и др., 1984; Lahiri et al., 1992]. Выделенную ДНК замораживали и хранили при температуре -20°С до проведения эксперимента.
Генотипирование осуществляли с помощью полимеразной цепной реакции (ПЦР), используя структуру праймеров и параметры температурных циклов, описанных в литературе (табл. 5). Смесь для ПЦР содержала 0,5-2,0 мкл специфической пары праймеров с концентрацией 1 о.е./мл, 1,2-1,8 мкл 10´ буфера для амплификации с концентрацией MgCl2 0,5-2,0 mM, 0,5-1,0 е.а. Taq ДНК-полимеразы («Сибэнзим», «Медиген», Новосибирск) и 100-200 нг геномной ДНК. Смесь помещали в 0,5 мл пробирки типа «Эппендорф», наслаивали сверху минеральное масло для предотвращения испарения и амплифицировали в автоматических минициклерах «MJ Rеsearch» (США) и «ЦиклоТемп 105» (Россия-Австрия).
Программа амплификации включала предварительную денатурацию при 94°С в течение 5 минут, с последующими 30-35 циклами отжига при температуре 60°С (1мин.), элонгации цепи при 72°С (40 сек.) и денатурации при 94°С (40 сек.). Программу завершала финальная элонгация при 72°С в течение 3 минут. Амплификат подвергали гидролизу соответствующей рестриктазой (табл.4) при оптимальной для фермента температуре на протяжении 12-24 ч. Рестрикционная смесь включала 5-7 мкл амплификата, 1,0-1,2 мкл 10´ буфера для рестрикции, поставляемого фирмой – производителем («Сибэнзим», Новосибирск), и 1-5 единиц активности фермента (в зависимости от эффективности его работы). Продукты рестрикции фракционировали 20-30 минут в 3% агарозном геле при напряжении 120 В. Фрагменты ДНК окрашивали бромистым этидием и визуализировали в ультрафиолетовом свете с применением компьютерной видеосъемки на приборе «UV-VIS Imager-II» (США).
Таблица 5
Характеристики исследованных полиморфных вариантов генов системы биотрансформации ксенобиотиков
Ген |
Локализация | Полиморфизм | Структура праймеров |
Фермент реакции |
Литературный источник |
| СYP2C19 |
10q24.1-24.3Экзон 5 |
681G>A |
5’-aat-tac-aac-cag-agc-ttg-gc 5’-tat-cac-ttt-cca-taa-aag-caa-g |
SmaI | De Morais et al., 1994 |
| CYP2E1 |
10q24.3-qter 5’-фланкирующий регион |
1293G>C |
5’-cca-gtc-gag-tct-aca-ttg-tca 5’-ttc-att-ctg-tct-tct-aac-tgg |
PstI | Salama et al., 1999 |
| 10q24.3-qter Интрон 6 | 7632T>A |
5’-ctg-ctg-cta-atg-gtc-act-tg 5’-gga-gtt-caa-gac-cag-cct-ac |
DraI | Lin et al., 1998 | |
| GSTM1 | 1p13.3 | Делеция |
5’-tgc-ttc-acg-tgt-tat-gga-ggt-tc 5’-gtt-ggg-ctc-aaa-tat-acg-gtg-g |
- | Spurdle et al., 2001 |
| GSTT1 | 22q11.23 | Делеция |
5’-ggt-cat-tct-gaa-ggc-caa-gg 5’-ttt-gtg-gac-tgc-tga-gga-cg |
- | |
| GSTP1 |
11q13 Экзон 5 |
313A>G |
5’- gta-gtt-tgc-cca-agg-tca-ag 5’- agc-cac-ctg-agg-ggt-aag |
BsoMAI | Ishii et al., 1999 |
2.2.3. Статистические методы анализа
Распределение генотипов по исследованным полиморфным локусам проверяли на соответствие равновесию Харди-Вайнберга (РХВ) с помощью точного теста Фишера [Вейр, 1995].
Ожидаемую гетерозиготность полиморфизма генов цитохромов Р450 (CYP2C19 и CYP2E1) и глутатионовых S-трансфераз (GSTT1, GSTM1 и GSTP1) рассчитывали по опубликованным методикам [Животовский, 1984]. Относительное отклонение ожидаемой гетерозиготности от наблюдаемой (D) рассчитывали по формуле:
D=(hobs–hexp)/hexp,
где hobs и hexp – ожидаемая и наблюдаемая гетерозиготность соответственно.
Для анализа ассоциации маркеров исследуемых генов с туберкулезом и бронхиальной астмой, а также с качественными патогенетически важными признаками заболеваний, сравнивали частоты аллелей и генотипов в группах больных и здоровых индивидов, используя критерий χ2 с поправкой Йетса на непрерывность, а также с применением двустороннего точного критерия Фишера.
Об ассоциации разных генотипов (или их комбинаций) с заболеваниями судили по величине отношения шансов (odds ratio (OR)) [Pearce, 1993], величины, показывающей, во сколько раз выше вероятность заболеть для индивида с определенным генотипом (или комбинацией генотипов):
OR= (A/B)/(C/D), где
А – число (процент) людей с данным генотипом (комбинацией генотипов) в группе больных;
С - число (процент) людей с данным генотипом (комбинацией генотипов) в группе здоровых;
В – число (процент) индивидов, не имеющих данного генотипа (комбинации генотипов) в группе больных;
D - число (процент) индивидов, не имеющих данного генотипа (комбинации генотипов) в группе здоровых.
Значения OR>1 указывают на возможную положительную ассоциацию с заболеванием. Обсуждение величин OR проводили при уровне значимости не более 5%.
При анализе семейного материала для поиска ассоциаций с генетическими маркерами был использован тест на неравновесие по сцеплению – Transmission/Disequilibrium Test (TDT):
TDT= (b-c)2/(b+c)2,
где b и c – число наследуемых аллелей от гетерозиготных родителей [Spielman, 1993]. TDT рассматривает вероятности передачи маркерного аллеля от гетерозиготного родителя больному потомку и отклонение этой вероятности от 0,5 может появиться только тогда, когда есть сцепление между маркерным локусом и локусом, контролирующим болезнь [Аксенович, 2001]. Этот тест имеет ряд существенных преимуществ: помимо того, что обладает высокой статистической мощью, к тому же устойчив к эффектам популяционной структуры (подразделенность, гетерогенность, примесь и т.д.), практически безотносителен к типу наследования.
При статистически значимых отклонениях распределения количественных признаков от нормального (по данным теста Шапиро-Уилки), сравнение проводили с помощью непараметрических критериев Манна-Уитни и Краскела-Уоллиса.
Для сравнения средних значений признаков до и после лечения применяли тест Уилкоксона [Гланц, 1998].
Для оценки параметров распределения, включая средние значения, стандартные отклонения использовали стандартный набор статистических процедур [Лакин, 1990]. Усреднение статистически значимых коэффициентов корреляции проводили с помощью z-преобразования Р. Фишера, после проверки на значимость различий полученных коэффициентов корреляции [Лильин и др., 1984].
Все расчеты проводили с использованием пакета прикладных программ “STATISTIСA for Widows 6.0” и в программе “Microsoft Excel 97”.
ГЛАВА 3. РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
3.1. Полиморфизм генов глутатионовых S-трансфераз (GSTT1, GSTM1, GSTP1) и цитохромов Р450 (CYP2E1, CYP2C19) у жителей г. Томска
Для реализации поставленных задач в рамках данного исследования проведен анализ полиморфизма генов биотрансформации ксенобиотиков I и II фазы метаболизма в отношении риска развития БА и ТБ, а также для значимых при заболеваниях качественных и количественных признаков.
В подавляющем большинстве случаев совместное функционирование обеих фаз метаболизма обеспечивает обезвреживание десятков тысяч ксенобиотиков всех химических классов. Предположительно, эта система возникла или эволюционировала в результате адаптации к техногенному загрязнению среды. Однако, учитывая, что загрязнение среды стало серьезным только в конце XX века, а также, что система метаболизма играет важную роль в перекисном окислении липидов и в биотрансформации многих эндогенных веществ, например, холестерина, витамина D, простагландинов и лейкотриенов, желчных кислот, токоферолов, стероидов и т.д. [Waxman, Azaroff, 1992], можно предположить, что ферменты биотрансформации первоначально функционировали как система метаболизма эндогенных веществ, а с изменяющимися условиями окружающей среды стали участвовать в метаболизме ксенобиотиков вследствие их широкой субстратной специфичности.
Одной из важных особенностей ферментативной системы метаболизма являются достаточно выраженные этнические различия. Это один из фактов, который необходимо учитывать при анализе связи полиморфизма генов с развитием заболевания, так как восприимчивость к патологии индивида одной популяции, определяющаяся сочетанием, как правило, распространенных вариантов генов, значительно различается с таковой в других популяционных группах. Поэтому, прежде чем анализировать данные гены в отношении риска возникновения болезни, следует рассмотреть популяционные особенности исследуемой группы здоровых индивидов г. Томска по данным локусам.
В ходе выполнения работы было исследовано 6 полиморфных вариантов генов цитохромов Р450 - CYP2C19 (681G>A) и CYP2E1 (7632T>A; 1293G>C) и глутатионовых S-трансфераз – GSTT1 (делеция), GSTM1 (делеция) и GSTP1 (313A>G).
Для изученных полиморфных вариантов генов GSTP1 (313A>G) и цитохромов Р450 - CYP2E1 (7632T>A; 1293G>C), CYP2C19 (681G>A) в контрольной выборке распределение генотипов соответствовало ожидаемым при равновесии Харди-Вайнберга (табл. 6). Для полиморфизма 313А>G гена GSTP1 отмечена незначительная гетерогенность между наблюдаемыми и ожидаемыми значениями генотипов за счет недостатка гетерозигот.
Осуществить проверку распределения генотипов по генам GSTT1 и GSTM1 на соответствие ожидаемым при равновесии Харди-Вайнберга невозможно, поскольку не устанавливалось гетерозиготного носительства.
Частоты встречаемости аллелей и генотипов изучаемых локусов в исследованной выборке здоровых жителей г. Томска оказались близки к значениям таковых в других европеоидных популяциях (рис. 2; табл. 6, 7). У жителей г. Томска частота «нулевого» генотипа для генов GSTT1 и GSTM1 составила 23,7% (n=32) и 54,8% (n=74), соответственно. Сходная ситуация описана для GSTT1 и GSTM1 описана для жителей г. Новосибирска [Ляхович и др., 2000; Вавилин и др., 2002]. Полученные частоты как для генов GSTT1 и GSTM1, так и для остальных полиморфных вариантов генов системы метаболизма, изучаемых в настоящей работе близки к значениям таковых в других европеоидных популяциях.
Таблица 6
Распределение генотипов по маркерам генов ферментов метаболизма ксенобиотиков у здоровых индивидов г. Томска
| Ген |
Полимор- физм |
Генотип | N.O. | N.E. | Частота аллеля |
c2 (d.f.=1) |
hobs hexp |
D |
| GSTP1 | 313A>G |
AA AG GG |
58 45 16 |
54,46 52,09 12,46 |
G=32,4 | 1,97 |
0,3782 0,4377 |
-0,136 |
| CYP2C19 | 681G>A |
*1/*1 *1/*2 *2/*2 |
90 26 0 |
91,46 23,09 1,46 |
CYP2C19*2=11,2 | 1,32 |
0,2241 0,1990 |
-0,126 |
| CYP2E1 | 7632T>A |
ТТ ТА АА |
106 20 2 |
105,13 21,75 1,13 |
A=9,4 | 0,45 |
0,1562 0,1699 |
-0,081 |
| CYP2E1 | 1293G>C |
C1C1 C1C2 C2C2 |
113 9 0 |
113,17 8,67 0,17 |
C2=3,7 | 0,04 |
0,0738 0,0710 |
-0,038 |
Примечание. N.O. и N.E. – наблюдаемая и ожидаемая численность генотипов; критерий c2 использован для оценки соответствия наблюдаемого распределения генотипов ожидаемому при равновесии Харди-Вайнберга; d.f. – число степеней свободы; hobs и hexp – наблюдаемая и ожидаемая гетерозиготность, соответственно; D –относительное
отклонение наблюдаемой гетерозиготности от ожидаемой.
Рис. 2. Частоты аллелей полиморфных вариантов генов GSTP1, CYP2E1 и CYP2C19 у жителей г. Томска (собственные данные) и в различных этнических группах (по: Brockmoller et al., 1996; Morita et al., 1997; Farker et al., 1998; Lin et al., 1998; Ishii et al., 1999; Miller et al., 2002; Yang et al., 2004).
Таблица 7
Частоты «нулевых» генотипов генов глутатионовых S-трансфераз в некоторых этнических группах
| Ген | Этническая принадлежность | Частота «нулевого» генотипа, % | Литературный источник |
| GSTT1 | Европеоиды США, n=152 | 17,1 | Crump et al., 2000 |
| Корейцы, n=220 | 45,9 | Kim et al., 2000 | |
| GSTM1 | Европеоиды CША, n=927 | 54,0 | Miller et al., 2002 |
| Корейцы, n=220 | 44,1 | Kim et al., 2000 |
Учитывая функциональную значимость системы метаболизма ксенобиотиков, многие работы по изучению полиморфизма генов соответствующих ферментов в первую очередь направлены на изучение эффективности лекарственной терапии различных заболеваний и связанными с ней проявлениями побочных эффектов. Другим аспектом интереса к этой системе является то, что в некоторых случаях можно оценить вклад в развитие заболевания полиморфизмов генов, отвечающих за взаимодействие организма человека с факторами окружающей среды. Кроме того, ферменты системы биотрансформации ксенобиотиков активно задействованы в метаболизме различных эндогенных веществ, в том числе и медиаторов воспаления. Поэтому можно предполагать, что наличие генетически обусловленных индивидуальных особенностей функционирования этой системы способствует развитию патологий. Глутатионовым S-трансферазам и цитохромам Р450 придают важное значение в формировании подверженности к заболеваниям, триггерами которых выступают неблагоприятные факторы внешней среды, например, онкологическая патология, инфекционные и аллергические заболевания и др. Из исследуемых генов наиболее изученными в отношении БА и ТБ являются гены глутатионовых S-трансфераз, причем касательно ТБ работы направлены в основном на изучение гепатотоксичности от применяемых для лечения заболевания препаратов, а не на поиск участия генов в развитии и патогенезе болезни.
В целом, результаты анализа частот аллелей и генотипов генов GSTT1, GSTM1, GSTP1, CYP2E1 и CYP2C19 у жителей г. Томска показывают значения близкие для европеоидных популяций, что закономерно, так как этническая принадлежность была одним из основных критериев отбора для исследования.
3.2. Оценка роли полиморфизма генов ферментов метаболизма ксенобиотиков в развитии бронхиальной астмы и туберкулеза
3.2.1. Ассоциация полиморфных вариантов генов GSTT1, GSTM1, GSTP1, CYP2E1 и CYP2C19 с атопической бронхиальной астмой
Интерес исследователей к изучению полиморфизма генов ферментов биотрансформации ксенобиотиков при БА, отвечающих за взаимодействие организма с факторами окружающей среды, вызван экологической обусловленностью заболевания. Показано, что «нулевые» генотипы генов GSTT1 и GSTM1 как в отдельности, так и в комбинации друг с другом являются факторами генетической предрасположенности к БА [Иващенко и др., 2001]. Кроме того, выявлена связь клинических особенностей течения заболевания с полиморфизмом генов GSTT1, GSTM1, CYP1A1 и NAT2 и показана модифицирующая роль курения на развитие БА [Ляхович и др., 2000, 2002]. Отмечены связь фенотипических проявлений БА (положительные кожные тесты, высокий уровень общего IgE) с полиморфизмом 313A>G гена GSTP1 ферментативной системы биотрансформации [Fryer et al., 2000] и различия в его функциональной значимости для БА атопического и неатопического генеза [Tamer et al, 2004]. При исследовании связи полиморфизма 313A>G гена GSTP1 с астмой, вызванной воздействием толуол диизоцианата установлено, что частота гомозиготного генотипа G/G этого гена снижена у индивидов с астмой, а также понижена у лиц, имеющих размах гиперреактивности бронхов от умеренной до критической по сравнению с индивидами, показывающими нормальную и низкую активность таковой по результатам диагностического теста с метахолином [Cristina et al., 2002]. Кроме того, показано, что материнское наследование G/G и A/A генотипов полиморфизма 313A>G гена GSTP1 может влиять на последующее развитие астмы и формирование особенностей ее клинического фенотипа у детей [Carroll et al., 2005].
Таблица 8
Распределение генотипов исследуемых генов ферментов биотрансформации у больных бронхиальной астмой и здоровых
| Генотип | БА | Контрольная группа | р | ||
| n | % | n | % | ||
| GSTT1 | |||||
| GSTT1+ | 99 | 75,6 | 103 | 76,3 | 1,000 |
| GSTT1 0/0 | 32 | 24,4 | 32 | 23,7 | |
| GSTM1 | |||||
| GSTM1+ | 38 | 29,0 | 61 | 45,2 | 0,008 |
| GSTM1 0/0 | 93 | 71,0 | 74 | 54,8 | |
| GSTP1 313A>G | |||||
| AA | 55 | 48,7 | 58 | 48,7 | 0,648 |
| AG | 47 | 41,6 | 45 | 37,8 | |
| GG | 11 | 9,7 | 16 | 13,5 | |
| CYP2E1 7632T>A | |||||
| TT | 88 | 72,1 | 106 | 82,8 | 0,049 |
| TA | 33 | 27,1 | 20 | 15,6 | |
| AA | 1 | 0,8 | 2 | 1,6 | |
| CYP2E1 1293G>C | |||||
| C1C1 | 106 | 89,8 | 113 | 92,6 | 0,498 |
| C1C2 | 12 | 10,2 | 9 | 7,4 | |
| C2C2 | 0 | 0,0 | 0 | 0,0 | |
| CYP2C19 681G>A | |||||
| *1/*1 | 91 | 66,4 | 90 | 77,6 | 0,068 |
| *1/*2 | 44 | 32,1 | 26 | 22,4 | |
| *2/*2 | 2 | 1,5 | 0 | 0,0 | |
Примечание. n – абсолютное значение; р – достигнутый уровень значимости по точному тесту Фишера.
По локусам GSTT1, GSTP1 и CYP2E1 (1293G>C) существенных различий между выборкой больных БА и контрольной группой не обнаружено (табл. 8). Для гена GSTM1 показано, что частота функционального генотипа встречалась у больных значимо реже (р=0,008). Это позволяет говорить о том, что для носителей делеции гена GSTM1, повлекшей за собой утрату активности соответствующего фермента, существует вероятность дисбаланса процессов детоксикации экзогенных и эндогенных веществ, что повышает для них риск развития заболевания. Об этом свидетельствует и тот факт, что носители делеционного генотипа показывают в два раза выше риск развития БА по сравнению с индивидами, имеющими функциональный генотип (OR=2,02; 95% CI: 1,18-3,46; p=0,009). Эта ассоциация может быть следствием множественности биологических функций глутатионовых S-трансфераз: участие в метаболизме эндогенных медиаторов воспаления (простагландинов, лейкотриена С4), нейромедиаторов. В настоящее время ведется дискуссия по поводу взаимосвязи ферментов метаболизма с регуляторным цитокиновым звеном. Предполагается, что образующиеся реактивные метаболиты в ходе реакций I фазы биотрансформации ксенобиотиков и их дальнейшее ковалентное связывание с макромолекулами клетки может привести к образованию аутоантигенов, вызывающих клеточный или гуморальный иммунный ответ [Ляхович и др., 2000]. Учитывая важную роль глутатионовых S-трансфераз в метаболизме эндогенных и экзогенных соединений, можно предполагать, что образующиеся реактивные метаболиты (даже при условии повышенного их образования в силу полноценной работы ферментов I фазы) у индивидов с функциональными генотипами GST, эффективно утилизируются, и, таким образом, не способствуют развертыванию воспалительных реакций и утяжелению уже начавшегося воспалительного процесса.
Исследование полиморфизма 7632T>A гена CYP2E1 показало, что гетерозиготный генотип T/A чаще встречался в группе больных БА (c2=4,22, р=0,049). Оценка OR показала, что гетерозиготные носители имеют повышенный риск развития заболевания по сравнению с контрольной группой (OR=2,0; 95% CI: 1,03-3,91; p=0,040). Данное обстоятельство является несколько неожиданным, поскольку предполагается, что для гетерозигот характерна более благоприятная активность соответствующего фермента для поддержания состояния здоровья, а не низкая или высокая у гомозигот, так как известно, что с аллелем 7632А часто ассоциированы редкие мутации, влияющие на каталитическую активность соответствующего белка [Hu et al., 1997].
При сравнении распределения частот генотипов полиморфизма 681G>A гена CYP2C19 фермента I фазы метаболизма показана гетерогенность между группами больных БА и здоровыми за счет тенденции к повышению частот гетерозигот *1/*2 у пробандов c БА (c2=3,32, р=0,068). При анализе частот аллелей этого локуса отмечена тенденция к преобладанию аллеля CYP2C19*2 у больных (c2=3,52, р=0,061) по сравнению со здоровыми.
Таблица 9
Распределение индивидов разного пола по группам, характеризующим степень тяжести бронхиальной астмы
| Группы сравнения | Легкая БА | Среднетяжелая БА | Тяжелая БА | |||
| n | % | n | % | n | % | |
| Мужчины | 14 | 56,0 | 32 | 60,4 | 19 | 36,5 |
| Женщины | 11 | 44,0 | 21 | 39,6 | 33 | 63,5 |
Примечание. n – абсолютное значение индивидов в группе.
Учитывая, что БА имеет довольно сложный клинический фенотип, проявляющийся от повышенной чувствительности бронхов до летальных форм, проследить участие генов системы биотрансформации в патогенезе возможно только при тщательном анализе особенностей сформировавшегося клинического фенотипа БА. Поэтому было проанализировано распределение частот генотипов и аллелей изучаемых локусов в отношении групп с различной степенью тяжести заболевания. Оценка тяжести БА основывается на клинических проявлениях, кроме того, учитывается вариабельность изменения бронхиальной проходимости в течение суток.
При сравнении групп, различающихся по степени тяжести (табл. 9) показано, что в группе с тяжелой БА преобладали индивиды женского пола (63,5%) в отличие от двух других представленных групп.
Рис. 3. Распределение «нулевых» (“0/0”) генотипов генов GSTT1 и GSTM1 у больных бронхиальной астмой, различающихся по степеням тяжести.
У больных БА с различными степенями тяжести заболевания установлены различия между легкой (n=26) и тяжелой (n=50) степенью тяжести по частотам генотипов гена GSTT1: «нулевой» генотип гена преобладал у лиц с легкой БА (38,5% и 16,0% у больных легкой и тяжелой БА, р=0,045) (рис. 3).
Предположив, что ферменты метаболизма ксенобиотиков способствуют сенсибилизации, осуществляя детоксикацию низкомолекулярных соединений, что клинически может проявляться проявлениями поливалентной аллергии, были проанализированы результаты кожных аллергопроб с полиморфизмом исследуемых генов ферментов метаболизма ксенобиотиков. В результате анализа не выявлено статистически значимых различий между группами, различающимися по наличию/отсутствию сенсибилизации (р>0,05).
В целом, полученные данные свидетельствуют о связи изученных полиморфных вариантов генов у жителей г. Томска как с БА, так и с отдельными клиническими признаками заболевания. Результаты работы показали вклад делеционного полиморфизма гена GSTM1 в развитии БА, основанного на функциональной значимости глутатионовых S-трансфераз в метаболизме эндогенных и экзогенных ксенобиотиков, а также выявили, что риск развития БА увеличивает гетерозиготный генотип гена CYP2E1 (полиморфизм 7632T>A). Кроме того, установлено значимое увеличение частоты «нулевого» генотипа гена GSTT1 у больных с легким течением заболевания.
3.2.2. Ассоциация полиморфизма генов ферментов метаболизма ксенобиотиков с туберкулезом
Известно, что при воспалении и инфекции происходит изменение уровня активности цитохромов Р450. Этот эффект опосредован цитокинами, которые угнетают транскрипцию генов и накопление мРНК различных изоформ цитохрома Р450 в клетках [Renton, 2004]. Такая реакция взаимодействия иммунной системы и системы метаболизма ксенобиотиков на внедрение инфекционного агента обеспечивает защиту от последствий возможной неконтролируемой активации потенциально опасной для организма ферментативной системы и снижает риск развития оксидативного стресса. В связи с этим представляется важным изучение полиморфизма генов системы метаболизма ксенобиотиков в отношении ТБ инфекции.
Таблица 10
Распределение генотипов исследуемых генов ферментов биотрансформации у больных туберкулезом и здоровых
| Генотип | ТБ | Контрольная группа | р | ||
| n | % | n | % | ||
| GSTT1 | |||||
| GSTT1 + | 254 | 81,9 | 103 | 76,3 | 0,195 |
| GSTT1 0/0 | 56 | 18,1 | 32 | 23,7 | |
| GSTM1 | |||||
| GSTM1 + | 126 | 40,6 | 61 | 45,2 | 0,404 |
| GSTM1 0/0 | 184 | 59,4 | 74 | 54,8 | |
| GSTP1 313A>G | |||||
| AA | 144 | 47,2 | 58 | 48,7 | 0,034 |
| AG | 142 | 46,6 | 45 | 37,8 | |
| GG | 19 | 6,2 | 16 | 13,5 | |
| CYP2E1 7632T>A | |||||
| ТТ | 246 | 80,4 | 106 | 82,8 | 0,235 |
| ТА | 59 | 19,3 | 20 | 15,6 | |
| АА | 1 | 0,3 | 2 | 1,6 | |
| CYP2E1 1293G>C | |||||
| C1C1 | 287 | 92,3 | 113 | 92,6 | 0,498 |
| C1C2 | 24 | 7,7 | 9 | 7,4 | |
| C2C2 | 0 | 0,0 | 0 | 0,0 | |
| CYP2C19 681G>A | |||||
| *1/*1 | 234 | 76,5 | 90 | 77,6 | 0,828 |
| *1/*2 | 69 | 22,5 | 26 | 22,4 | |
| *2/*2 | 3 | 1,0 | 0 | 0,0 | |
Примечание. n – абсолютное значение; р достигнутый уровень значимости по точному тесту Фишера.
При сравнении распределения частот аллелей и генотипов полиморфных вариантов генов метаболизма ксенобиотиков между группой больных ТБ и контрольной выборкой получены различия только для гена GSTP1, обусловленные увеличением доли гомозигот G/G у здоровых (р=0,034) (табл. 10). Установлена протективная роль генотипа G/G гена GSTP1 в отношении развития ТБ (OR=0,43; 95%CI: 0,20-0,91; р=0,026). В результате однонуклеотидной замены аденина на гуанин в этом гене происходит изменение ферментативной активности, обусловливающее повышение уровня гидрофобных аддуктов в тканях легких. Показано, что соответствующая замена изолейцина на валин в 105 положении, расположенная в субстрат-связывающем Н-участке фермента, приводит к неоднозначным изменениям кинетических параметров ферментам. Ген GSTP1 в большей степени экспрессируется в респираторном тракте. Возможно, что для гомозигот G/G гена GSTP1 характерна высокая каталитическая активность по отношению к ряду соединений, являющихся факторами риска развития ТБ (например, продукты табачного дыма), которая способствует их быстрому метаболизму и дальнейшему выведению, таким образом, снижая риск развития заболевания.
Сравнительная оценка распределения частот аллелей и генотипов генов GSTT1, GSTM1, CYP2E1 и CYP2C19 между группами больных ТБ и здоровых индивидов cтатистически значимых различий не показала (табл. 10).
Проведен анализ полиморфизма исследуемых генов для определения связи с отдельными клиническими проявлениями ТБ, характеризующими тяжесть и течение патологического процесса. Особенно важной является оценка связи исследуемых полиморфизмов с лабораторными показателями, свидетельствующими о степени тяжести туберкулезного процесса и выраженности воспалительной реакции.
Исходя из данных рентгенологического исследования, были сформированы три группы больных ТБ в зависимости от объема поражения легкого (I группа – поражено 1-2 сегмента легкого (n=76), II группа – поражена одна доля легкого (n=40), III группа – задействовано более доли легкого (n=117)). При анализе распределения частот аллелей и генотипов исследуемых локусов получены различия для гена CYP2C19: отмечено преобладание гомозигот *1/*1 в III-й группе по сравнению с I-ой (80,3% и 65,8%, соответственно, р=0,040) и повышенная частота аллеля CYP2C19*1 в III-й группе (р=0,045). Эти данные свидетельствуют в пользу гипотезы о накоплении в клетках активных форм кислорода и реактивных метаболитов, образующихся в реакциях I фазы. Известно, что CYP2C19 семейства цитохрома Р450 является основным ферментом, катализирующим в организме человека трансформацию (S)-мефенитоина в соответствующее 4’-гидроксипроизводное [Крынецкий, 1996]. Замена 681G>A в пятом экзоне гена является основным генетическим дефектом, приводящим к инактивации CYP2C19, проявляющимся на уровне фенотипа наличием медленных метаболизеров (S)-мефенитоина. Полученные различия в отношении развития ТБ можно трактовать следующим образом: у носителей CYP2C19*1 аллеля при метаболизме субстратов для CYP2C19 повышается уровень реактивных окислителей, которые независимо от работы ферментов II фазы, накапливаются в организме и оказывают повреждающее действие на клетки, усиливая патологический процесс. Развитие окислительного стресса при ТБ способствует усилению процессов деструкции в легочной ткани. Подобные результаты были получены при исследовании инсерционного полиморфизма гена CYP2E1 с развитием инфильтративного ТБ у жителей Башкортостана [Бикмаева и др., 2004].
Основной этиологической причиной в развитии туберкулезного процесса являются микобактерии ТБ, несмотря на важное значение факторов, способствующих развитию заболевания (курение, асоциальный образ жизни и др.). С момента открытия Р.Кохом в 1882 году, возбудитель ТБ достаточно полно изучен в отношении биологических свойств, характера и условия заражения. Сложилось четкое представление о патогенезе заболевания, его клинических проявлениях, течении и исходах. ТБ отличается клиническим полиморфизмом, что проявляется в развитии различных форм заболевания – от малых с бессимптомным течением до обширных деструктивных процессов с выраженной клинической картиной. Известно, что контакт организма человека с M. tuberculosis не обязательно приводит к развитию болезни [Хоменко, 1990]. В легких процесс начинается с формирования небольшого очага, где происходит накопление серозного экссудата с нейтрофильными гранулоцитами и макрофагами, подвергающегося первичному некрозу. В дальнейшем экссудативные изменения заканчиваются формированием туберкулезной гранулемы, затем при благоприятном течении процесс ограничивается рубцеванием или инкапсуляцией. Развитие туберкулезных изменений в ранее не инфицированном M. tuberculosis организме свидетельствует о первичном генезе заболевания. В случае обострения существующего первичного процесса, либо повторного экзогенного инфицирования, возникает вторичный ТБ [Цинзерлинг, 1996]. Несмотря на накопленные данные о патогенезе заболевания до сих пор не решен вопрос, почему в одних случаях развивается очаговый ТБ, а в других – инфильтративный или другие клинические формы [Хоменко, 1990].
Предположив, что различия в патогенезе заболевания связаны с индивидуальными особенностями генома человека, был изучен полиморфизм генов системы метаболизма в группах больных ТБ, различающихся по клиническим формам заболевания. Наиболее характерной клинической формой заболевания среди исследуемых больных жителей г. Томска является инфильтративный ТБ (50,3%) (табл. 11). По Сибирскому Федеральному округу также отмечается преобладание вторичного ТБ, среди которых инфильтративная форма занимает первое место в общей структуре клинических форм и составляет 55,1% [Краснов, 2004].
Процентное соотношении доли индивидов мужского и женского пола колеблется в зависимости от клинической формы заболевания. В выборке больных ТБ г. Томска отмечена высокая заболеваемость диссеминированной формой у мужчин по сравнению с женщинами (р=0,047), для остальных форм статистически значимых различий не показано (р=0,602-0,783). По мнению многих исследователей, более высокая заболеваемость у мужчин обусловлена не только биологическими свойствами макроорганизма, но и особенностями условий труда и влиянием различных вредных привычек [Рабухин, 1976; Хоменко, 1990].
Таблица 11
Половозрастной состав групп больных туберкулезом с различными клиническими формами
| Группы сравнения | Мужчины n, (%) |
Женщины n, (%) |
Cуммарно n, (%) | Средний возраст ± S.D. |
| ТБ внутригрудных лимфоузлов | 21 (9,9) | 13 (12,6) | 34 (10,8) | 6,8 ± 3,4 |
| Очаговый ТБ | 17 (8,1) | 10 (9,7) | 27 (8,6) | 35,7 ± 14,1 |
| Диссеминированный ТБ | 53 (25,1) | 15 (14,6) | 68 (21,7) | 36,9 ± 13,3 |
| Инфильтративный ТБ | 104 (49,3) | 54 (52,4) | 158 (50,3) | 33,4 ± 12,7 |
| Остальные формы | 16 (7,6) | 11 (10,7) | 27 (8,6) | 36,4 ± 11,6 |
Примечание. n – численность в группе; S.D. – стандартное отклонение.
При сравнительном изучении распределения частот «нулевого» генотипа гена GSTM1 группы больных первичным ТБ с поражением внутригрудных лимфоузлов и пациентов с инфильтративной формой отмечена тенденция к снижению доли лиц, гомозиготных по делеции, среди индивидов с ТБ вторичного генеза (70,6% и 52,3%, соответственно, р=0,058).
Анализ частот генотипов полиморфизма 313A>G гена GSTP1 в группах больных с ТБ внутригрудных лимфоузлов и инфильтративным ТБ также показал различия между этими двумя группами (р=0,026) за счет преобладания гетерозигот A/G у лиц больными вторичным ТБ по сравнению с первичной формой (50,0% и 34,4% соответственно).
Таким образом, отмечена связь полиморфных вариантов генов ферментов I и II фаз метаболизма GSTP1 313A>G и CYP2C19 681G>A в развитии ТБ легких у русских жителей г. Томска. Показана протективная роль генотипа G/G полиморфизма гена GSTP1 313A>G для развития ТБ легких. В то же время, установлено возможное модифицирующее влияние полиморфизма 681G>A гена CYP2C19 на увеличение объема зоны поражения легочной ткани при уже возникшем заболевании. Эти данные говорят о том, что изученные полиморфные варианты генов системы метаболизма оказывают вклад на развитие туберкулезного процесса у русских г. Томска.
3.2.3. Сравнительный анализ роли полиморфных вариантов генов системы метаболизма ксенобиотиков в детерминации бронхиальной астмы и туберкулеза
Согласно поставленной задаче, в ходе исследования был проведен сравнительный анализ распределения частот аллелей и генотипов полиморфных вариантов исследуемых локусов между группами больных ТБ и БА. Для делеционного полиморфизма гена GSTM1 показано, что частота «нулевого» генотипа у больных БА превышает таковую в группе больных ТБ (71,0% и 59,4% соответственно; c2=4,85, р=0,028) (рис. 4), а относительный риск развития БА по сравнению с ТБ у носителей «нулевого» генотипа гена GSTM1 составил 1,68 (95% CI: 1,06-2,67). Полученные данные еще раз подтверждают высокую важность функционирования глутатионовой S-трансферазы m1 в развитии БА, поскольку были получены различия в частотах генотипов данного полиморфизма гена GSTM1 при сравнении больных БА и контрольной группы (р=0,008). Для полиморфных вариантов генов глутатионовых S-трансфераз q1 и p1 при сравнении частот генотипов статистически значимых различий не показано (р=0,162 и р=0,387 соответственно).
|
|
|
Рис. 4. Частоты «нулевых» (0/0) генотипов гена GSTM1 у больных бронхиальной астмой и туберкулезом.
При сравнении частот генотипов исследуемых в настоящей работе полиморфных вариантов генов цитохромов Р450 различий не было показано между группами больных БА и ТБ (р=0,079-0,437). Среди больных ТБ преобладал *1/*1 генотип полиморфизма 681G>A гена СYP2C19 по сравнению с больными БА (76,5% и 66,4% соответственно), однако эти различия статистически не значимы (р=0,079). В то же время были получены различия между группами больных с ТБ и БА при сравнении частот аллелей этого локуса, где частота СYP2C19*1 аллеля у больных ТБ выше по сравнению с больными БА (87,7% и 82,5% соответственно; c2=3,96, р=0,047). Интересно, что СYP2C19*1 аллель участвует в увеличении объема поражения легочной ткани у больных ТБ легких, и, по-видимому, данная ассоциация предполагает важное участие гена СYP2C19 именно в формировании клинических проявлений при ТБ. Соответственно, для индивидов носителей СYP2C19*2 аллеля среди больных БА относительный риск выше по сравнению с больными ТБ (OR=1,52, 95% CI: 1,01-2,30; p=0,047).
Таким образом, при сравнительном анализе распределения частот аллелей и генотипов полиморфных вариантов генов ферментов метаболизма ксенобиотиков между больными ТБ и БА были получены следующие различия: «нулевой» генотип гена GSTM1 и носительство СYP2C19*2 аллеля полиморфизма 681G>A гена СYP2C19 определяют развитие БА.
3.3. Анализ ассоциаций генов ферментов метаболизма ксенобиотиков с бронхиальной астмой и туберкулезом на семейном материале
Известно, что при анализе ассоциаций генетических факторов с болезнями и признаками по принципу «случай-контроль», вероятность ложноположительного результата высока, в связи с тем, что кроме возможной истиной значимости исследуемого гена в отношении изучаемой патологии необходимо учесть возможное неравновесие по сцеплению с другими генами, имеющими непосредственное отношение к болезни. Кроме того, крайне необходимо принять во внимание процессы, происходящие непосредственно при формировании популяции, например, подразделенность, метисация, инбридинг.
Поэтому, для исключения ложноположительной ассоциации с подверженностью к полигенным заболеваниям, наиболее перспективны являются исследования на семейном материале, которые позволяют исключить влияние факторов подразделенности популяции.
В связи с этим для анализа ассоциаций исследуемых полиморфных вариантов генов ферментов метаболизма ксенобиотиков в настоящем исследовании использовали тест на неравновесие при наследовании TDT (Transmission/Disequilibrium Test). Применение данного теста для диаллельного локуса позволяет сравнить частоту аллелей у больных потомков гетерозиготных родителей и, в случае, если один из аллелей будет встречаться чаще у пробандов, можно говорить об ассоциации с заболеванием [Spielman, 1993].
Таблица 12
Численность аллелей, унаследованных больными бронхиальной астмой и туберкулезом от гетерозиготных родителей
| Группы сравнения | Ген (полиморфизм) | |||
| GSTP1 (313A>G) |
CYP2E1 (7632T>A) |
CYP2C19 (681G>A) | ||
| БА | N гетерозиготных родителей | 38 | - | 39 |
| N унаследованных аллелей | A=13; G=25 | - | CYP2C19*1=24; CYP2C19*2=15 | |
| TDT (p) | 3,79 (0,052) | - | 2,08 (0,150) | |
| ТБ | N гетерозиготных родителей | 17 | 11 | - |
| N унаследованных аллелей | A=10; G=7 | Т=8; А=3 | - | |
| TDT (р) | 0,53 (0,467) | 2,30 (0,129) | - | |
Примечание. TDT – значение теста Transmission/Disequilibrium Test; р – достигнутый уровень значимости.
При анализе семейного материала больных БА наблюдалось предпочтительное наследование аллеля 313G гена GSTP1 больными от гетерозиготных родителей (TDT=3,79, р=0,052), близким к статистической значимости (табл. 12). Ген GSTP1 локализован на хромосоме 11q13, а для этого региона показано сцепление с бронхиальной гиперреактивностью и атопией [Daniels et al., 1996; Thomas et al., 1997]. Полученные данные позволяют предполагать возможное участие глутатионовых S-трансфераз p1 в детоксикации и элиминации токсических продуктов в эпителиальных тканях респираторного тракта. Данные о неоднозначных изменениях каталитической активности при мутации 105Val [Watson et al., 1998], позволяют предположить, что недостаток соответствующего фермента, задействованного в метаболизме ксенобиотиков, приводит к нарушению детоксикации электрофильных реактивных метаболитов, образующихся в I-й фазе биотрансформации и оказывающих повреждающее действие на бронхи, тем самым, провоцируя развитие БА у предрасположенных индивидов.
Для гена CYP2E1 (7632T>A) не удалось проследить наследование аллелей для больных БА в силу низкой гетерозиготности родителей пробандов. Использование TDT для гена CYP2C19 (681G>A) не показало значимой ассоциации с заболеванием (TDT=2,08, p=0,150). Значение TDT для полиморфизма 313A>G гена GSTP1 не показало преимущественного наследования ни одного из аллелей пробандами больными ТБ (TDT=0,53, p=0,467), в отличие от такового значения для больных БА. В случае полиморфизма по «нулевым» аллелям для генов GSTT1 и GSTM1 применение TDT затруднено по причине невозможности определения гетерозиготного носительства.
В целом, семейный анализ наследования аллелей генов ферментов метаболизма ксенобиотиков показал отсутствие предпочтительного наследования аллелей больными потомками от гетерозиготных родителей, однако для аллеля 313G гена GSTP1 получено значение TDT близкое к статистической значимости.
3.4. Оценка связи комбинаций генотипов генов ферментов биотрансформации ксенобиотиков с туберкулезом и бронхиальной астмой
Спектр изоформ определяет соотношение метаболических путей биотрансформации ксенобиотиков и спектр образуемых метаболитов. В результате генетически обусловленного полиморфизма этих ферментов может возникать дефицит либо значительная активность отдельных изоформ, что определяет риск развития заболевания. Знания о физиологической функции ферментов метаболизма ксенобиотиков, свидетельствующие о четкой и скоординированной работе I и II фаз биотрансформации, позволяют предположить, что наиболее важная информация об их роли в патогенезе заболеваний будет получена при анализе носителей определенных сочетаний генотипов.
Поэтому, кроме анализа ассоциаций БА и ТБ с отдельными полиморфными вариантами генов GSTT1, GSTM1, GSTP1, CYP2E1 и CYP2C19 в данном исследовании были также изучены ассоциации различных комбинаций генотипов с заболеваниями (табл. 13, 14).
Комбинации генотипов GSTM1+ и GSTT1+, GSTT1+ и CYP2C19 *1/*1, GSTM1+ и CYP2E1 Т/Т, CYP2C19*1/*1 и CYP2E1 C1/C1 можно охарактеризовать, как генетические факторы устойчивости к возникновению БА. Объяснением резистентности к патологии для носителей сочетания функциональных генотипов GSTM1+/GSTT1+, может быть их функциональная значимость в отношении всей системы метаболизма, способствующая своевременной утилизации ксенобиотиков, оказывающих вредное воздействие на организм, следствием которого может быть увеличение риска развития БА. Протективное сочетание генотипов генов ферментов I фазы CYP2C19 и CYP2E1, позволяет предположить участие в метаболизме возможных триггеров БА, исходя из знаний об их функции в организме. Во всех остальных случаях протективная роль комбинаций генотипов в отношении БА показана при сочетании аллелей генов, которые обеспечивают нормальное функционирование соответствующих ферментов системы метаболизма обеих фаз. В отношении ТБ протективную роль показали следующие комбинации генотипов: GSTM1 0/0 и CYP2E1 Т/А, GSTP1 G/G и CYP2E1 Т/Т, GSTP1 G/G и CYP2E1 C1/C1 (табл. 13).
Таблица 13
Протективные комбинации генотипов в отношении развития бронхиальной астмы и туберкулеза
| Комбинация генотипов | БА | ТБ | ||
| OR (95% CI) | р | OR (95% СI) | р | |
| GSTM1+ и GSTP1 G/G |
0,10 (0,0-0,76) |
0,018 |
0,37 (0,14-0,98) |
0,045 |
| GSTM1+ и GSTT1+ |
0,47 (0,26-0,84) |
0,009 | - | - |
| GSTT1+ и CYP2C19 *1/*1 |
0,51 (0,29-0,88) |
0,014 | - | - |
| GSTM1+ и CYP2E1 Т/Т |
0,28 (0,14-0,56) |
0,000 | - | - |
| CYP2C19*1/*1 и CYP2E1 C1/C1 |
0,53 (0,29-0,95) |
0,032 | - | - |
| GSTM 0/0 и CYP2E1 Т/А | - | - |
0,15 (0,06-0,42) |
0,000 |
| GSTP1 G/G и CYP2E1 Т/Т | - | - |
0,35 (0,16-0,79) |
0,009 |
| GSTP1 G/G и CYP2E1 C1/C1 | - | - |
0,39 (0,18-0,85) |
0,015 |
Примечание. OR – значение отношения шансов; 95% CI – 95% доверительный интервал; р достигнутый уровень значимости по точному тесту Фишера.
Отмечена «общая» комбинация генотипов GSTM1+ и GSTP1 G/G, оказывающая протективную роль как в отношении развития БА (OR=0,10; 95% CI: 0,0-0,76; p=0,018), так и ТБ (OR=0,37; 95% CI: 0,14-0,98; p=0,045) (табл. 13). Несмотря на то, что согласно TDT, отмечено предпочтительное наследование аллеля 313G гена GSTP1 у больных БА, гомозиготы по этому аллелю в сочетании с функциональным генотипом гена GSTM1 показывают устойчивость к развитию заболевания. Возможно, что наличие особенностей ферментов метаболизма, таких как множественность форм и перекрывающаяся субстратная специфичность, позволяют существенно восполнить дефекты индивидуального фермента в метаболизме ксенобиотиков активностью других.
При анализе комбинаций генотипов полиморфных вариантов генов ферментов метаболизма ксенобиотиков установлены сочетания генотипов, предрасполагающие к развитию БА (табл. 14).
Таблица 14
Комбинации генотипов, предрасполагающие к развитию бронхиальной астмы
| Комбинация генотипов | OR (95% CI) | р |
| GSTТ1+ и GSTМ1 0/0 | 1,89 (1,13-3,19) | 0,015 |
| GSTM1 0/0 и CYP2E1 Т/А | 3,18 (1,31-7,87) | 0,008 |
Примечание. OR – значение отношения шансов; 95% CI – 95% доверительный интервал; р достигнутый уровень значимости по точному тесту Фишера.
Выявлена комбинация генотипов GSTM1 0/0 и CYP2E1 Т/А, являющаяся фактором риска развития БА (OR=3,18; 95% CI: 1,31-7,87; р=0,008). В данном случае можно предположить наличие взаимосвязанной регуляции между двумя соответствующими ферментами глутатионовой S-трансфераза и цитохрома Р450. По этому поводу имеются данные о скоординированной экспрессии GSTM1 и GSTM3 в легочной ткани человека [Anttila et al., 1995], а также о более высокой активности CYP1A2 у индивидуумов с нулевым генотипом GSTM1 [MacLeod et al., 1997]. Стоит отметить, что эта же комбинация генотипов показала в отношении развития ТБ протективное значение (OR=0,15; 95% CI: 0,06-0,42; р=0,000). Видимо такая комбинация определяет неэффективный/эффективный метаболизм различных триггеров БА и ТБ эндогенного и экзогенного происхождения.
Возможно, высокий риск для носителей комбинации генотипов GSTТ1+ и GSTМ1 0/0 (OR=1,89; 95% CI: 1,13-3,19, р=0,015) получен вследствие высокой важности для данного заболевания GSTM1, и даже наличие функционального генотипа GSTT1 не снижает риск развития заболевания. Можно предположить, что соответствующие ферменты могут также метаболизировать различные по химической структуре молекулы, тогда именно субстраты для GSTМ1 могут быть триггерами БА и присутствие функционального генотипа GSTT1 никаким образом не оказывает влияния на сохранение состояния здоровья.
Среди всех проанализированных комбинаций полиморфных вариантов генов не показано ни одного сочетания, имеющего патогенетическую значимость в развитии ТБ.
В заключение следует сказать, что при сравнении сочетаний генотипов генов ферментов I и II фаз метаболизма для различных по этиопатогенезу заболеваний отмечена общая протективная комбинация генотипов GSTM1+ и GSTP1 G/G в развитии ТБ и БА. Для ТБ не показано ни одного патогенетически значимого сочетания генотипов генов ферментов метаболизма ксенобиотиков. Кроме того, комбинация генотипов GSTM1 0/0 и CYP2E1 Т/А являющаяся фактором риска развития БА, для ТБ оказывает протективную роль.
3.5. Связь полиморфизма генов ферментов метаболизма ксенобиотиков с изменчивостью количественных признаков у больных бронхиальной астмой и туберкулезом
Следующим этапом настоящего исследования было изучение связи исследуемых полиморфных вариантов генов с изменчивостью значимых для заболеваний количественных признаков, характеризующей адаптационные способности организма. Возможно, что изучаемые полиморфизмы цитохрома Р450 и глутатионовых S-трансфераз имеют значение в развитии БА и ТБ в целом, а также в выраженности отдельных клинических проявлений. Поэтому представлялось важным оценить наличие связи исследуемых генов с количественными лабораторными показателями, характеризующими особенности течения анализируемых заболеваний.
Известно, что ключевой особенностью БА является состояние бронхиальной гиперреактивности, свидетельствующее о повышенном бронхоконстрикторном ответе на различные физико-химические факторы, когда бронхоспазм развивается в ответ на воздействие, не вызывающее такой реакции у большинства здоровых лиц. На этом основан клинический тест с метахолином, показывающий изменения чувствительности и реактивности бронхов.
Предположив, что индивидуальная способность к детоксикации веществ, способствующих развитию БА и бронхиальной гиперреактивности, детерминирована полиморфизмом генов системы метаболизма ксенобиотиков, были проанализированы значения дозы метахолина (по результатам теста на бронхиальную гиперреактивность) с изученными полиморфными вариантами исследуемых генов. Признак не показал корреляции с возрастом обследуемых (r=-0,359, p=0,066). Учитывая значимые отклонения уровня метахолина от закона Гаусса (по данным теста Шапиро-Уилки, W=0,782, p=0,001), сравнение было проведено с помощью непараметрического медианного теста. В результате была показано близкое к статистически значимому различие «количественного фенотипа» БА у мужчин с полиморфизмом 313A>G гена GSTP1: для гомозиготных носителей GG генотипа характерна более низкая доза метахолина, по сравнению с мужчинами-носителями АА и AG генотипов (рис. 5, табл. 15). Следует отметить, что в доступных нам литературных источниках отмечается связь аллеля 313А гена GSTP1 с бронхиальной гиперреактивностью для европеоидной популяции [Cristina et al., 2002].
Известно, что основными соединениями, вызывающими бронхиальную гиперреактивность, являются реактивные окислители ключевые компоненты воспалительной реакции. Бронхиальная гиперреактивность может быть модулирована уровнем реактивных окислителей, возможно, с помощью их способности регулировать продукцию эйкозаноидов через стимуляцию освобождения арахидоновой кислоты.
Таблица 15
Взаимосвязь изменчивости уровня метахолина с распределением генотипов полиморфизма гена GSTP1 313A>G
| Генотип |
|
|
|
|||
| Мужчины (n=25) | ||||||
| AA | 4,000 | 6,261 | -2,261 | |||
| AG | 11,000 | 8,348 | 2,652 | |||
| GG | 1,000 | 1,391 | -0,391 | |||
| p=0,054* | ||||||
| Женщины (n=16) | ||||||
| AA | 5,000 | 5,625 | -0,625 | |||
| AG | 2,000 | 1,875 | 0,125 | |||
| GG | 3,000 | 2,500 | 0,500 | |||
| p=0,789* | ||||||
Примечание. ![]() obs – наблюдаемые средние значения,
obs – наблюдаемые средние значения, ![]() exp
ожидаемые средние значения,
exp
ожидаемые средние значения, ![]() =
=![]() obs-
obs-![]() exp, * – достигнутый уровень значимости медианным тестом.
exp, * – достигнутый уровень значимости медианным тестом.
Гены глутатионовых S-трансфераз являются генами-кандидатами для одного из клинических проявлений астмы – бронхиальной гиперреактивности, а, следовательно, и для БА, поскольку кодируемые ими ферменты понижают уровень реактивных окислителей [Hayes, McLellan, 1999]. Эта точка зрения подтверждается исследованиями, показавшими, что индивиды с пониженной антиоксидантной способностью имеют повышенный риск атопической БА и уменьшение потока антиоксидантов ассоциировано с экспрессией связанных с астмой фенотипов.
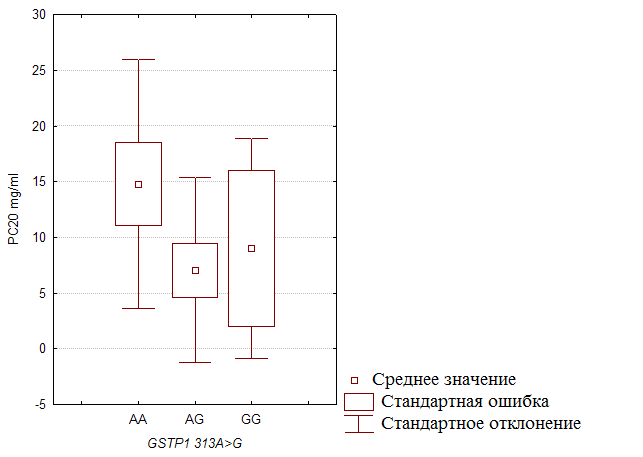
Рис. 5. Уровни метахолина у носителей различных генотипов полиморфизма 313A>G гена GSTP1 у лиц мужского пола.
Полиморфизм в генах GSTT1 и GSTM1 не показал связи с бронхиальной гиперреактивностью, что может отражать различия в генной экспресии, также как изменчивости в метаболизме субстратов, имеющих отношение для развития БА. Действительно, несмотря на то, что в эпителиальных клетках легких человека экспрессируются различные генные продукты GST, глутатионовые S-трансферазы p класса составляют более чем 90% от общей GST-активности [Frayer et al., 1986].
Известно, что для БА аллергического характера характерно значительное повышение уровня общего IgE. Через IgE-опосредованный механизм целый ряд клеточных элементов: тучные клетки, макрофаги, лимфоциты, эпителиальные и эндотелиальные клетки независимо друг от друга или совместно принимают участие в воспалении дыхательных путей, тем самым, осуществляя иммунный ответ организма на внедрение антигена. В этом контексте была рассмотрена гипотеза, предполагающая зависимость изменчивости уровня общего IgE от генетического полиморфизма ферментов метаболизма ксенобиотиков. Было показано значимое повышение уровня IgЕ у женщин с генотипом *1/*1 гена CYP2C19 по сравнению с носителями остальных генотипов (табл. 16).
Таблица 16
Распределение уровня IgE у носителей различных генотипов гена CYP2C19 (полиморфизма 681G>A) среди женщин
| Генотип | n | Средние значения IgE±S.E. | p |
| *1/*1 | 20 | 408,0±73,4 | 0,044 |
| *1/*2+*2/*2 | 4 | 67,5±26,7 |
Примечание. n – абсолютное значение человек в группе; р – достигнутый уровень значимости для теста Манна-Уитни.
Анализ изменчивости уровня общего IgE у больных БА с другими, изученными в данной работе полиморфными вариантами генов метаболизма ксенобиотиков, не показал ассоциаций ни у мужчин, ни у женщин (р>0,05).
Таблица 17
Значение показателей спирометрии ± S.E. в зависимости от генотипа по полиморфизмам генов глутатионовых S-трансфераз GSTT1 и GSTM1 у больных бронхиальной астмой
| Группа сравнения | Генотип | Форсированная жизненная емкость легких | Объем форсированного выдоха за 1 секунду | Пиковая скорость выдоха |
| GSTT1 | ||||
| Мужчины |
GSTT1 + (n=19) |
2,32±0,41 | 1,97±0,45 | 4,31±1,11 |
|
GSTT1 0/0 (n=12) |
2,58±0,50 | 2,29±0,42 | 4,82±0,84 | |
| р* | 0,122 | 0,059 | 0,187 | |
| Женщины |
GSTT1 + (n=18) |
2,46±0,73 | 2,02±0,67 | 3,86±1,33 |
|
GSTT1 0/0 (n=5) |
2,42±0,34 | 2,09±0,70 | 4,26±1,18 | |
| р* | 0,911 | 0,831 | 0,550 | |
| GSTM1 | ||||
| Мужчины |
GSTM1 + (n=8) |
2,31±0,42 | 1,92±0,31 | 4,31±1,08 |
|
GSTM1 0/0 (n=23) |
2,46±0,47 | 2,15±0,50 | 4,58±1,08 | |
| р* | 0,415 | 0,236 | 0,538 | |
| Женщины |
GSTM1 + (n=4) |
1,78±0,58 | 1,64±0,50 | 2,97±1,31 |
|
GSTM1 0/0 (n=19) |
2,59±0,59 | 2,12±0,67 | 4,15±1,22 | |
| р* | 0,021 | 0,191 | 0,097 | |
Примечание. n – объемы выборок; * - уровень значимости для однофакторного дисперсионного анализа.
Учитывая важность показателей исследования функции внешнего дыхания у больных БА для оценки степени тяжести заболевания, проведен сравнительный анализ связи полиморфных вариантов генов метаболизма с основными спирометрическими показатели: форсированная жизненная емкость легких (FVC), объем форсированного выдоха за 1 секунду (FEV1) и пиковая скорость выдоха (PEF). Отмечена связь FVC с полиморфизмом гена GSTM1 среди женщин (F=6,263, p=0,021), у мужчин таких различий не наблюдается (табл. 17). Кроме того, показаны близкая к статистической значимости связь FEV1 с полиморфизмом гена GSTT1 у мужчин, а также PEF с полиморфизмом гена GSTM1 у женщин.
Учитывая, что патогенные свойства M. tuberculosis в условиях развивающегося специфического процесса в легких непосредственно сказываются на особенностях реагирования системы крови, для больных ТБ были проанализированы параметры общего анализа крови: уровень гемоглобина, количество эритроцитов, лейкоцитов, скорость оседания эритроцитов (СОЭ); а также параметры биохимического анализа крови: количество билирубина (прямой и обратный), аланинаминотрансферазы (АЛТ) до начала лечения и через два месяца после лечения.
Воздействие инфекционного агента вызывает развитие комплекса изменений как специального, так и стрессового характера. Последние оказывают непосредственное влияние на формирование основного патологического процесса, в то же время специфика развивающегося туберкулезного процесса определяет особенности реакций общего адаптационного синдрома.
Для полиморфных вариантов генов глутатионовых S-трансфераз не показано связи с изменчивостью количественных показателей периферической крови (табл. 18). Однако получена ассоциация полиморфизма 681G>A гена CYP2C19 фермента I-й фазы метаболизма ксенобиотиков у мужчин: аллель CYP2C19*2 связан с низким уровнем эритроцитов (р=0,027) (табл. 19), для них также отмечена тенденция к снижению уровня гемоглобина (р=0,065). Для женщин таких различий не показано (р>0,05). Известно, что при ТБ имеет место снижение количества эритроцитов как за счет их ускоренного разрушения в периферической крови под влиянием токсических фракций M. tuberculosis, так и вследствие нарушения эритропоэза в результате туберкулезной интоксикации [Глебович, 1951; Милосердова, 1958; Шмелев, 1959; Радзинский, 1961; Кан, 1972].
Таблица 18
Средние значения (±S.E.) количественных параметров крови больных туберкулезом носителей разных генотипов полиморфизмов генов глутатионовых S-трансфераз
| Группа сравнения | Генотип |
Гемоглобин (г/л) |
Эритроциты (х1012/л) |
Лейкоциты (х109/л) |
СОЭ (мм/ч) |
| 1 | 2 | 3 | 4 | 5 | 6 |
| GSTT1 | |||||
| Мужчины | GSTT1 + |
130,01±1,78 n=115 |
4,17±0,06 n=112 |
8,23±0,33 n=116 |
25,14±1,76 n=115 |
| GSTT1 0/0 |
135,35±2,43 n=26 |
4,29±0,10 n=25 |
8,01±0,50 n=28 |
25,59±3,36 n=28 |
|
| p | 0,251* | 0,353* | 0,892* | 0,486* | |
| Женщины | GSTT1 + |
119,14±1,04 n=69 |
3,90±0,06 n=65 |
7,06±0,36 n=69 |
25,41±2,26 n=67 |
| GSTT1 0/0 |
121,82±1,99 n=13 |
3,72±0,14 n=12 |
7,04±0,51 n=13 |
23,94±6,96 n=12 |
|
| p | 0,613** | 0,322* | 0,489* | 0,739* | |
| GSTM1 | |||||
| Мужчины | GSTM1 + |
129,26±2,42 n=51 |
4,10±0,09 n=49 |
8,56±0,45 n=53 |
27,50±2,61 n=52 |
| GSTM1 0/0 |
131,98±1,96 n=90 |
4,25±0,06 n=88 |
7,96±0,36 n=91 |
23,00±1,92 n=91 |
|
| p | 0,263* | 0,091* | 0,158* | 0,205* | |
| Женщины | GSTM1 + |
120,66±3,32 n=29 |
3,94±0,10 n=27 |
6,93±0,56 n=29 |
26,27±3,74 n=28 |
| GSTM1 0/0 |
118,97±2,36 n=53 |
3,83±0,06 n=50 |
7,12±0,39 n=53 |
24,58±2,69 n=51 |
|
| p | 0,676** | 0,393* | 0,491* | 0,656* | |
| GSTP1 313A>G | |||||
| Мужчины | AA |
132,72±2,07 n=76 |
4,23±0,07 n=76 |
8,48±0,39 n=79 |
24,51±1,92 n=79 |
| AG+GG |
128,85±2,33 n=62 |
4,14±0,07 n=62 |
7,91±0,42 n=62 |
24,56±2,66 n=61 |
|
| p | 0,201* | 0,309* | 0,521* | 0,871* | |
| Женщины | AA |
119,88±2,81 n=34 |
3,97±0,07 n=31 |
7,35±0,56 n=34 |
24,39±3,89 n=33 |
| AG+GG |
119,69±2,65 n=47 |
3,80±0,08 n=45 |
6,85±0,38 n=47 |
25,94±3,00 n=47 |
|
| р | 0,512* | 0,207* | 0,670* | 0,461* | |
Примечание. В скобках указаны единицы измерения; n объемы выборок;
*- достигнутый уровень значимости теста Манна-Уитни; ** - уровень значимости для однофакторного дисперсионного анализа.
Таблица 19
Средние значения (±S.E.) количественных параметров крови у больных туберкулезом носителей разных генотипов полиморфных вариантов генов цитохромов Р450
| Группа сравнения | Генотип |
Гемоглобин (г/л) |
Эритроциты (х1012/л) |
Лейкоциты (х109/л) |
СОЭ (мм/ч) |
|
| 1 | 2 | 3 | 4 | 5 | 6 | |
| CYP2E1 7632T>A | ||||||
| Мужчины | ТТ |
132,11±1,80 n=105 |
4,24±0,06 n=103 |
7,98±0,32 n=108 |
24,19±1,80 n=107 |
|
| ТА+АА |
127,94±2,98 n=34 |
4,05±0,08 n=32 |
8,99±0,57 n=34 |
25,75±3,25 n=34 |
||
| p | 0,204* | 0,212* | 0,091* | 0,686* | ||
| Женщины | ТТ |
120,07±2,20 n=67 |
3,88±0,06 n=62 |
7,06±0,36 n=67 |
26,31±2,46 n=65 |
|
| ТА+АА |
116,84±3,95 n=14 |
3,81±0,11 n=14 |
7,243±0,638 n=14 |
20,31±4,65 n=13 |
||
| p | 0,532** | 0,707* | 0,549* | 0,283* | ||
| CYP2E1 1293G>C | ||||||
| Мужчины | C1C1 |
131,29±1,62 n=122 |
4,21±0,05 n=119 |
7,93±0,28 n=125 |
25,28±1,68 n=124 |
|
| C1C2 |
129,11±4,50 n=19 |
4,10±0,15 n=18 |
9,84±1,04 n=19 |
20,46±3,90 n=19 |
||
| p | 0,925 | 0,669 | 0,060 | 0,215 | ||
| Женщины | C1C1 |
119,42±1,96 n=80 |
3,87±0,05 n=75 |
7,04±0,32 n=80 |
25,92±2,20 n=77 |
|
| C1C2 |
119,00±5,20 n=3 |
0,23±0,13 n=3 |
6,83±1,24 n=3 |
14,09±11,68 n=3 |
||
| p | 0,968** | 0,845* | 0,855* | 0,219* | ||
| CYP2C19 681G>A | ||||||
| Мужчины | *1/*1 |
133,09±1,68 n=102 |
4,27±0,06 n=99 |
8,45±0,34 n=104 |
23,85±1,75 n=104 |
|
|
*1/*2+ *2/*2 |
124,81±3,40 n=36 |
3,96±0,08 n=35 |
7,60±0,52 n=37 |
26,66±3,52 n=36 |
|
|
| p | 0,065* | 0,027* | 0,195* | 0,477* |
|
|
| Женщины | *1/*1 |
121,04±2,17 n=57 |
3,90±0,06 n=53 |
7,20±0,35 n=57 |
25,37±2,90 n=57 |
|
|
*1/*2+ *2/*2 |
115,89±3,98 n=24 |
3,78±0,10 n=23 |
6,83±0,69 n=24 |
25,57±4,04 n=23 |
|
|
| р | 0,469** | 0,469* | 0,341* | 0,795* |
|
|
Примечание. В скобках указаны единицы измерения; n – объемы выборок;
*- достигнутый уровень значимости теста Манна-Уитни; ** - уровень значимости для однофакторного дисперсионного анализа.
Данные об экспрессии гена CYP2C19 в костном мозге, позволяют предполагать, что наличие аллеля CYP2C19*2 приводит к снижению функции соответствующего фермента, поэтому у индивидов, носителей мутантного аллеля течение ТБ может сопровождаться разрушающим действием токсинов M. tuberculosis на клетки костного мозга, что приводит к неэффективному эритропоэзу. Подобное предположение о связи делеционного полиморфизма гена GSTT1, сопровождающимся отсутствием соответствующего фермента II-й фазы биотрансформации ксенобиотиков, с неспособностью метаболизировать токсичные для гемопоэтических клеток субстраты, нашло свое подтверждение в исследовании о развитии приобретенной апластической анемии у детей [Dirksen et al., 2004].
Анализ остальных параметров периферической крови: лейкоцитов и скорости оседания эритроцитов не показал влияния исследуемых в работе полиморфизмов генов ФМК на изменчивость вышеперечисленных показателей как для мужчин, так и для женщин. Однако отмечена тенденция к повышению уровня лейкоцитов у носителей гетерозиготных генотипов полиморфных вариантов генов CYP2E1 7632T>A и CYP2E1 1293G>C среди мужчин (р=0,091, р=0,060 соответственно). Учитывая низкую частоту аллелей этих полиморфных вариантов, можно предположить, что статистическая мощность исследованной выборки оказалась недостаточной, чтобы установить значимую связь в отношении изменения уровня лейкоцитов крови.
Метаболизм лекарственных препаратов и эффекты их дальнейшего пребывания в организме в большей степени зависят от генетического полиморфизма ферментов системы биотрансформации. На сегодня известно, что человек имеет 59 активных генов семейства цитохрома Р450, и 6 из них кодируют важные для лекарственного метаболизма ферменты [Ingelman-Sundberg, 2004]. Как отмечалось ранее, для ферментов биотрансформации характерна способность к метаболизму большого количества субстратов по причине того, что ферменты I-й и II-й фаз биотрансформации перекрываются в своей субстратной специфичности. Однако для многих форм Р450 выделены специфические лекарства, используемые для фармакокинетических оценок. Для исследуемых в настоящей работе цитохромов Р450 и глутатионовых S-трансфераз селективные субстраты представлены в табл. 20.
Основным органом, участвующим в метаболизме лекарств, является печень, где обозначены самые высокие концентрации ферментов метаболизма по сравнению с другими органами и наибольшее разнообразие экспрессируемых форм [Райс, Гуляева, 2000]. Полиморфизм генов метаболизма ксенобиотиков в настоящее время активно изучается в отношении индивидуальной чувствительности к лекарственной терапии и, особенно в проявлении многообразных побочных реакций, связанных с лечением.
Таблица 20
Специфичные субстраты для ферментов системы метаболизма
| Фермент | Специфичный субстрат | Литературный источник |
| CYP2E1 | Хлорзоксазон | Kharasch et al., 1993 |
| CYP2C19 | S-мефенитоин | De Morais et al., 1994 |
| GSTT1 | Трансстильбеноксид | Hallier et al., 1993 |
| GSTM1 | Хлористый метилен и хлористый метил | Seidegard et al., 1988 |
| GSTP1 | Этакриновая кислота и бензпирендиолэпоксид | Awasthi et al., 1993 |
Показано, что при биотрансформации новокаинамид превращается в метаболит, который может вызвать у медленных ацетиляторов картину болезни, похожую на красную волчанку, а сульфазалин у этих людей может вызвать лейкопению, гепатотоксичность и нейропатии. Эффективность терапии ТБ зависит как от индивидуальных способностей индивида в метаболизме лекарств, так и от взаимоотношения антимикобактериальных препаратов с системой цитохромов Р450 непосредственно самой микобактерии ТБ. Обнаружено, что геном M. tuberculosis содержит гены, кодирующие 20 различных цитохромов Р450, в том числе ферментов, являющихся мишенью действия для противогрибковых препаратов. Кроме того, опубликованные данные об угнетении метаболизирующей функции печени за счет снижения содержания цитохромов Р450 в этом органе при бактериальной и вирусной инфекции через цитокин-опосредованные механизмы, позволяют предполагать изменение фармакодинамики, а соответственно токсичности лекарственных препаратов [Prandota, 2002].
Во всех странах получило признание комбинированное применение химиопрепаратов, позволяющее добиться бактерицидного эффекта и предотвратить развитие лекарственной устойчивости в процессе лечения. Принцип комбинированного применения нескольких химиопрепаратов известен давно, еще в 1955 г. он был внедрен в практику химиотерапии как метод предупреждения лекарственной устойчивости M. tuberculosis. Актуальность лекарственных поражений печени во фтизиатрии обусловлена необходимостью полихимиотерапии туберкулеза, что создает высокую медикаментозную нагрузку на больного, и в большей степени ее испытывает печень, осуществляя метаболизм туберкулостатиков и патогенетических средств. Противотуберкулезные препараты изониазид, рифампицин, пиразинамид обладают значительной гепатотоксичностью (особенно этот эффект выражен при их комбинации), этамбутол, микобутин и другие – в меньшей степени. Лекарственные гепатиты у больных туберкулезом относят к категории преимущественно токсических побочных реакций химиотерапии.
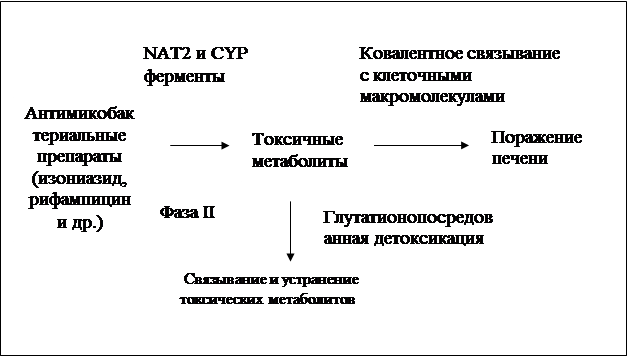
Рис. 6. Взаимодействие между лекарственными препаратами и ферментативной системой метаболизма ксенобиотиков, приводящее к лекарственно-индуцированному гепатиту (по: Roy et al., 2001).
Частым осложнением при лечении ТБ легких производными гидразина изоникотиновой кислоты, например, изониазидом, являются гепатотоксические реакции. Известно, что чаще они возникают у лиц, быстро инактивирующих изониазид, поскольку у них высвобождается значительно больше гидразина, в частности, ацетилгидразина, который может вызывать дистрофические поражения печени (рис. 6).
Аланинаминотрансфераза (АЛТ) — фермент, катализирующий трансаминирование, присутствует во многих тканях организма, в частности, в печени. В гепатоцитах он локализуется главным образом в цитозольной фракции.
Высвобождение АЛТ в кровь происходит при нарушениях внутренней структуры гепатоцитов и повышении проницаемости клеточных мембран, что свойственно как острому вирусному гепатиту, так и рецидивам хронического гепатита. В этой связи АЛТ считается индикаторным ферментом, и к его определению прибегают постоянно при постановке диагноза гепатитов любой природы.
Установлено статистически значимое увеличение уровней АЛТ (р=0,001) и билирубина (р=0,05) после двух месяцев применения антимикобактериальных препаратов (табл. 21). Значение АЛТ не показало корреляции с возрастом и полом (r=-0,161 и r=-0,152, соответственно, р>0,05). Выявлена ассоциация полиморфного варианта 313A>G гена глутатионовой S-трансферазы p1 (GSTP1) с увеличением активности АЛТ после лечения противотуберкулезными препаратами в течение двух месяцев (р=0,021) (табл. 22).
Поскольку метаболизм изониазида и рифампицина приводит к образованию более токсичных метаболитов, то одной из возможных причин полученного различия может быть прямая связь между генотипом индивида и изменением уровня активности показателя печеночной функции. Такой факт закономерен, так как известно, что глутатионовые S-трансферазы играют значительную роль в метаболизме противотуберкулезных препаратов, таких как изониазид и рифампицин [Sodhi et al. 1996; Sodhi et al., 1997].
Таблица 21
Изменения уровней аланинаминотрансферазы и билирубина до и после двух месяцев лечения
| Значение уровня аланинаминотрансферазы (ммоль/(ч.л)) |
Значение уровня билирубина (мкмоль/л) |
||
|
До начала лечения |
После 2-х месяцев лечения |
До начала лечения |
После 2-х месяцев лечения |
| 0,03-1,55 | 0,03-1,83 | 4,5-102,0 | 5,0-342,0 |
| 0,001 | 0,050 | ||
Примечание. В скобках указаны единицы измерения; р достигнутый уровень значимости для теста Уилкоксона.
В доступных источниках литературы показано, что рифампицин индуцирует экспрессию глутатионовых S-трансфераз, а изониазид-индуцированные повреждения печеночных клеток у модельных животных показывают связь с истощением содержащегося в печени глутатиона, и соответственно, с пониженной активностью GST. Эти эффекты максимальны, когда применяются два препарата совместно [Steele et al., 1991].
Полученные результаты представляют интерес в связи с тем, что последнее время появляются данные о развитии гепатотоксичности во время применения антимикобактериальных препаратов у лиц с определенным генотипом по генам ФМК. Так, показана связь «нулевого» генотипа гена GSTM1 с лекарственно-индуцированной гепатотоксичностью в Индии [Roy et al., 2001]. Исследования у 318 пациентов при лечении ТБ в Тайвани показали ассоциации полиморфизма СYP2E1 (Rsa I) с токсическим поражением печени [Huang et al., 2003].
Таблица 22
Средние уровни аланинаминотрансферазы и билирубина после двух месяцев лечения у носителей разных генотипов по генам глутатионовых S-трансфераз и цитохромов Р450 больных туберкулезом
|
Ген полиморфизм |
Генотип (n) | АЛТ±S.E. | р | Билирубин±S.E. | р |
|
GSTT1 del |
GSTT1 + (92) | 0,27±0,03 | 0,383* | 15,53±3,91 | 0,682* |
| GSTT1 0/0 (27) | 0,22±0,04 | 9,40±0,99 | |||
|
GSTM1 del |
GSTM1 + (51) | 0,20±0,02 | 0,149* | 10,81±1,89 | 0,557* |
| GSTM1 0/0 (68) | 0,29±0,04 | 16,63±5,12 | |||
|
GSTP1 313A>G |
AA (61) | 0,29±0,04 | 0,021** | 16,87±5,66 | 0,604** |
| AG (49) | 0,20±0,03 | 10,98±1,95 | |||
| GG (8) | 0,32±0,05 | 13,69±6,39 | |||
| CYP2C19 681G>A | *1/*1 (86) | 0,26±0,03 | 0,580* | 15,37±4,16 | 0,543* |
| *1/*2+ *2/*2 (32) | 0,22±0,02 | 10,74±1,62 | |||
| CYP2E1 7632T>A | TT (90) | 0,27±0,03 | 0,706* | 15,52±4,01 | 0,198* |
| TA+AA (29) | 0,21±0,03 | 9,84±0,68 | |||
| CYP2E1 1293G>C | C1C1 (106) | 0,26±0,03 | 0,976* | 14,66±3,41 | 0,603* |
| C1C2+C2C2 (13) | 0,22±0,04 | 9,85±1,18 |
Примечание. АЛТ±S.E.– средние значения уровня аланинаминотрансферазы со стандартной ошибкой; билирубин±S.E. – средние значения уровня билирубина со стандартной ошибкой; n – объем выборки; * - достигнутый уровень значимости по тесту Манна-Уитни; ** - достигнутый уровень значимости по тесту Краскела-Уоллиса.
Отмечена ассоциация статуса медленного ацетилятора NAT2 и гепатита, вызванного применением антимикобактериальных препаратов [Huang et al., 2002]. Данные проведенного исследования у жителей г. Томска предполагают участие полиморфного варианта гена GSTP1 (313A>G) в изменчивости уровня показателя печеночной функции при лечении ТБ антимикобактериальными препаратами.
Таким образом, в большинстве случаев для исследуемых количественных признаков наблюдали статистически значимые отклонения распределения от нормального (по данным теста Шапиро-Уилки). С учетом этого сравнение проводили с помощью непараметрических критериев Манна-Уитни, Краскела-Уоллиса и медианного теста. При анализе «количественного фенотипа» больных БА с распределением полиморфных вариантов генов системы метаболизма отмечено: тенденция к снижению уровня метахолина, вызывающего бронхоспазм для гомозиготных носителей аллеля 313G гена GSTP1, значимое повышение уровня IgE у носителей генотипа *1/*1 гена CYP2C19, связь делеционного полимофизма гена GSTM1 с изменчивостью показателя FVC, а также близкая к статистической значимости значения FEV1 и PEF c полиморфизмом генов глутатионовых S-трансфераз GSTM1 и GSTT1.
Оценка «количественного фенотипа» ТБ показала связь гена фермента I-ой фазы метаболизма CYP2C19 (полиморфизм 681G>A) с изменчивостью уровня эритроцитов. Анализ изменчивости показателей печеночной функции показал значимые различия в уровне АЛТ и билирубина до и после двух месяцев лечения антимикобактериальными препаратами. В ходе анализа выявлена ассоциация полиморфного варианта 313A>G гена глутатионовой S-трансферазы p1 (GSTP1) c увеличением уровня АЛТ.
ЗАКЛЮЧЕНИЕ
Ферментативная система метаболизма ксенобиотиков является практически универсальным механизмом, поддерживающим внутренний баланс и способствующим сохранности здоровья организма человека. Существовавшая изначально для метаболизма эндогенных субстратов, система эволюционировала, адаптируясь к техногенному загрязнению окружающей среды. В её функционировании задействованы уникальные по своим свойствам ферменты: гемопротеид – цитохром Р450, низкомолекулярный трипептид - глутатион и др. С помощью целых семейств этих ферментов с одинаковой каталитической активностью и различной субстратной специфичностью метаболизируются сотни самых разных по химическому составу соединений. Одним из важнейших свойств системы метаболизма является индукция – активация транскрипции гена в присутствии субстрата. Тканеспецифичная экспрессия различных изоформ метаболизма определяет ее адаптацию к структурно-функциональной организации той или иной системы организма. Наибольшая экспрессия ферментов в печени обеспечивает наиболее активное участие этого органа в метаболизме ксенобиотиков. В совокупности все ферменты, участвующие в деградации молекул ксенобиотиков, функционируют как единый, четко скоординированный комплекс. Поэтому отклонение их функции неизменно приводит к вредным для организма человека последствиям. Это обстоятельство подтверждают многочисленные исследования о функционировании системы метаболизма при различных воздействиях окружающей среды и патологических состояниях [Lin et al., 1998; Иващенко и др., 2000; Ляхович и др., 2000, 2002; Delfino et al., 2000; Вавилин и др., 2002; Rollinson et al., 2003; Бикмаева и др., 2004].
Согласно современным представлениям БА и ТБ относятся к группе дистропных болезней. Однако многочисленные проведенные исследования поиска генетической компоненты подверженности к этим заболеваниям показали ряд «общих» генов, белковые продукты которых задействованы на всех этапах патогенеза. С этой точки зрения целесообразным и перспективным представлялся сравнительный анализ полиморфных вариантов генов системы метаболизма ксенобиотиков, поскольку кодируемые ими ферменты задействованы в деградации эндогенных субстратов, а именно многочисленных медиаторов воспаления (простагландинов, лейкотриенов и т. д.), что легло в основу настоящего исследования.
Ряд работ показал связь генов ферментов метаболизма ксенобиотиков с развитием БА и её клиническими проявлениями в различных популяциях [Luszawaka-Kutrzela, 1999; Ляхович и др., 2000, 2002; Fryer et al., 2000; Иващенко и др., 2001; Gawronska-Szklarz et al., 2001; Вавилин и др., 2002; Gilliand et al., 2002; Сафронова и др., 2003; Brasch-Andersen et al., 2004; Tamer et al., 2004; Carroll, 2005]. Однако, учитывая значительные этнические различия в полиморфизме генов этой системы, существует противоречивая информация об их значимости для развития заболевания.
При оценке роли полиморфизма генов метаболизма ксенобиотиков для развития БА у жителей г. Томска показана ассоциация полиморфизма генов ферментов как I-й – CYP2E1, так и II фазы – GSTM1 с заболеванием.
Для носителей делеции гена GSTM1, приводящей к утрате активности соответствующего фермента, существует возможность дисбаланса процессов детоксикации экзогенных и эндогенных веществ, что повышает для них в два раза риск развития заболевания БА по сравнению с индивидами, имеющими функциональный генотип. Следует отметить, что подобные данные были получены во многих исследованиях, как для европеоидных, так и для монголоидных популяций [Вавилин и др., 2002; Ляхович и др., 2000; Zhang et al., 2004]. Можно предполагать, что эта ассоциация является важным следствием множественности биологических функций глутатионовых S-трансфераз и обусловлена их участием в метаболизме эндогенных медиаторов воспаления (простагландинов Н2, E2, F2a, лейкотриена С4). Однако интересно, что в проявлении тяжести заболевания не отмечена значимость этого гена, а у пациентов с легкой степенью тяжести преобладал делеционный генотип гена GSTT1. Тяжесть БА определяется многими факторами (пол, возраст начала, отягощенная наследственность, предшествующее лечение, сопутствующие аллергические заболевания), и в настоящее время нет четких представлений о формировании клинического полиморфизма заболевания [Огородова и др., 2002]. В данном случае, можно лишь предполагать, что при наличии отчетливо неблагоприятного генотипа, развитие патологического процесса может сдерживаться присутствием в геноме индивида генов, контролирующих выработку белковых структур, которые препятствуют развитию более тяжелой степени течения БА.
В ходе исследования были получены данные о связи ТБ с другими генами системы метаболизма. Так, в отношении инфекционного заболевания показана протективная роль полиморфиза 313A>G гена GSTP1 фермента II-й фазы метаболизма. Эта ассоциация объясняется с позиции высокой экспрессии глутатионовой S-трансферазы π1 в легких, защищающих таким образом человека на пути воздействия на организм токсичных агентов окружающей среды (например, химических соединений, содержащихся в табачном дыме и выхлопных газах), которые можно отнести к факторам, провоцирующим развитие ТБ.
Анализ полиморфизма исследуемых генов в формировании и степени выраженности клинических проявлений ТБ показал, что последствия возможной активации CYP2C19 нарушают оксидантное равновесие при уже развившемся заболевании, а развитие окислительного стресса способствует усилению процессов деструкции в легочной ткани.
Несомненно, единственной причиной развития ТБ является инфицирование организма M. tuberculosis. Однако дальнейшая судьба возбудителя болезни зависит от многих факторов, которые в совокупности определяют полиморфизм клинических форм заболевания. Так, показаны различия между группами больных с ТБ внутригрудных лимфоузлов и инфильтративным ТБ для полиморфизма 313A>G гена GSTP1, играющего роль в подверженности к заболеванию.
В исследованиях дизайна «случай-контроль» особую важность приобретает использование для анализа ассоциаций генетических факторов с заболеванием семейного материала, позволяющего проследить наследование аллелей, связанных с болезнью. В ходе данного исследования показано предпочтительное наследование аллеля 313G гена GSTP1 больными БА потомками от гетерозиготных родителей.
Важная информация о взаимодействии ферментов системы метаболизма двух фаз для оценки их вклада в подверженность к заболеваниям была получена при анализе носителей определенных сочетаний генотипов. Отмечена комбинация генотипов генов ферментов II фазы метаболизма GSTM1 и GSTP1, оказывающая протективную роль как в отношении развития БА, так и ТБ. В большинстве случаев протективная роль комбинаций генотипов в отношении БА показана при сочетании аллелей генов, которые обеспечивают полноценное функционирование соответствующих ферментов системы метаболизма обеих фаз. Выявлена комбинация генотипов полиморфных вариантов генов GSTM1 и CYP2E1, предрасполагающая к развитию БА, но оказывающая протективную роль в отношении ТБ. Среди всех проанализированных комбинаций полиморфных вариантов генов не показано ни одного сочетания, имеющего патогенетическую значимость в развитии ТБ. Полученные данные свидетельствуют, что эффекты комбинаций определенных генотипов генов ФМК различны в развитии БА и ТБ.
Следующим этапом исследования было изучение связи исследуемых полиморфных вариантов генов с количественными лабораторными показателями, характеризующими особенности течения различных по этиологии и патогенезу заболеваний. Учитывая варьирование количественных признаков в зависимости от пола, оценка вклада полиморфизма генов системы биотрансформации ксенобиотиков была проведена отдельно для мужчин и женщин и показала участие генов ферментов как I-й так и II-й фаз метаболизма. Так отмечена связь гена CYP2C19 с изменчивостью IgE у женщин и GSTM1 – с показателем форсированной жизненной емкости легких, которые относятся к важным количественным характеристикам проявлений БА. Оценка гематологических показателей крови у мужчин, больных ТБ выявила связь полиморфизма гена CYP2C19 с изменчивостью уровня эритроцитов в периферической крови. Неодинаковый характер ассоциаций генов ферментов метаболизма с количественными признаками у мужчин и женщин позволяет предположить, что та часть структуры наследственной компоненты предрасположенности к заболеваниям, которая связана с полиморфизмом этих генов, неодинакова у представителей разного пола, что выражается в дифференциальной частоте многих болезней у мужчин и женщин в одной популяции.
Особую ценность для практического здравоохранения приобретают результаты настоящего исследования в свете участия генов ферментов метаболизма ксенобиотиков в формировании гепатотоксичных реакций на противотуберкулезную терапию. Выявленная ассоциация повышения активности аланинаминотрансферазы после лечения антимикобактериальными препаратами с полиморфным вариантом гена GSTP1 в дальнейшем может использоваться для разработки комплекса профилактических мер по предотвращению побочных реакций от химиотерапии ТБ.
В целом, полученные результаты свидетельствуют, что наличие определенных генотипов и их комбинаций генов ферментов метаболизма ксенобиотиков может оказывать существенное влияние на предрасположенность и формирование клинического фенотипа БА и ТБ. Сравнительный анализ участия генов ферментов биотрансформации ксенобиотиков в развитии БА и ТБ позволил раскрыть некоторые генетические аспекты этих дистропных заболеваний. В ходе исследования показана дифференциация генов, задействованных в формировании клинического фенотипа заболеваний: гены GSTM1 и CYP2E1 связаны с БА и ее клиническими проявлениями, а GSTP1 - с развитием ТБ. Из исследуемых полиморфных вариантов генов ферментативной системы биотрансформации отмечен «общий» ген – CYP2C19, ассоциированный с изменчивостью признаков, характеризующих некоторые особенности течения этих двух заболеваний. Одним из предполагаемых функциональных механизмов, лежащих в основе полученных ассоциаций, может быть участие белковых продуктов соответствующих генов в метаболизме эндогенных ксенобиотиков, в том числе многочисленных медиаторов воспалительных реакций. Актуальность продолжения исследований сравнительного характера клинически различных групп заболеваний не вызывает сомнения, поскольку полученные результаты позволяют не только приблизиться к пониманию молекулярно-генетических основ подверженности к ним, но и в дальнейшем открывают перспективы профилактики их развития.
ВЫВОДЫ:
1. Исследованная выборка русских жителей города Томска по частотам аллелей и генотипов полиморфных вариантов генов цитохромов Р450 - CYP2C19 (681G>A), CYP2E1 (7632T>A; 1293G>C) и глутатионовых S-трансфераз – GSTT1 (делеция), GSTM1 (делеция), GSTP1 (313A>G) соответствует таковым для европеоидных популяций.
2. Риск развития бронхиальной астмы увеличивают «нулевой» генотип гена GSTM1 и гетерозиготный генотип по полиморфизму 7632T>A гена CYP2E1. Генотип G/G полиморфизма 313A>G гена GSTP1 снижает риск развития туберкулеза (OR=0,43; 95%CI: 0,20-0,91; p=0,026).
3. «Нулевой» генотип гена GSTT1 (р=0,045) выступает в качестве фактора, определяющего легкое течение бронхиальной астмы. Для полиморфного варианта 313A>G гена GSTP1 установлена ассоциация с инфильтративной формой туберкулеза (р=0,026); гомозиготный генотип *1/*1 полиморфизма 681G>A гена CYP2C19 преобладал у больных с распространенным процессом в легочной ткани (р=0,040).
4. Аллельные варианты генов ферментов метаболизма ксенобиотиков ассоциированы с «количественными фенотипами» болезней: для больных бронхиальной астмой отмечены связь полиморфизма 681G>A гена CYP2C19 с изменчивостью уровня IgE (р=0,044), делеционного полимофизма гена GSTM1 с изменчивостью форсированной жизненной емкости легких (p=0,021) у женщин; у мужчин, больных туберкулезом полиморфизм гена CYP2C19 связан с некоторыми гематологическими показателями – в частности, с уровнем эритроцитов (р=0,027).
5. Протективное значение имеет комбинация генотипов GSTM1+ и GSTP1 G/G в развитии бронхиальной астмы (OR=0,10; 95% CI: 0,0-0,76; p=0,018) и туберкулеза (OR=0,37; 95% CI: 0,14-0,98; p=0,045). Подверженность к астме увеличивают комбинации генотипов GSTT1+ и GSTM1 0/0 (OR=1,89; 95% CI: 1,13-3,19; р=0,015) и GSTM1 0/0 и CYP2E1 T/A (OR=3,18; 95% CI: 1,31-7,87; р=0,008), генотипическая комбинация GSTM1 0/0 и CYP2E1 T/A обеспечивает резистентность к туберкулезу (OR=0,15; 95% CI: 0,06-0,42; р=0,000).
6. У больных туберкулезом легких показано статистически значимое увеличение уровня показателей печеночной функции при применении антимикобактериальных препаратов. Установлена связь полиморфизма 313A>G гена GSTP1 с изменчивостью уровня аланинаминотрансферазы (р=0,021).
7. Выявлены различия в структуре генетической подверженности к бронхиальной астме и туберкулезу по генам ферментативной системы метаболизма ксенобиотиков: гены GSTM1, CYP2E1 и CYP2C19 связаны с бронхиальной астмой и значимыми для заболевания качественными и количественными признаками, а GSTP1 и CYP2C19 ассоциированы с туберкулезом и клиническими проявлениями инфекционной патологии.
ЛИТЕРАТУРА
1. Авербах М. М. Иммунология и иммунопатология туберкулеза. — М.: Медицина, 1976. - 311 с.
2. Аксенович Т.И Статистические методы генетического анализа признаков человека: Учеб. Пособие / Новосиб. гос. ун-т. Новосибирск, 2001. - 128 с.
3. Афанасьева И.С., Спицин В.А. Наследственный полиморфизм глутатион S-трансферазы печени человека в норме и при алкогольном гепатите // Генетика. – 1990. – Т. 26 (7). – С. 1309-1314.
4. Баранов В.С., Баранова Е.В., Иващенко Т.Э. и др. Геном человека и гены «предрасположенности». (Введение в предиктивную медицину).- СПб.: Интермедика, 2000.- 272 с.
5. Бикмаева А.Р., Сибиряк С.В., Хуснутдинова Э.К. Инсерционный полиморфизм гена CYP2E1 у больных инфильтративным туберкулезом легких в популяциях республики Башкортостан // Молекулярная биология. – 2004. – Т. 38. -№ 2. – С. 239-243.
6. Бикмаева А.Р., Сибиряк С.В., Хуснутдинова Э.К. Инсерционный полиморфизм гена CYP2E1 у больных инфильтративным туберкулезом легких и в популяциях республики Башкортостан // Молекулярная биология. – 2004. – Т. 38. - 2. – С. 239-243.
7. Бочков Н.П., Захаров А.Ф., Иванов В.И. Медицинская гентика. – М.: Медицина, 1984. – 366 с.
8. Вейр Б. Анализ генетических данных: Пер. с англ. – М.: Мир, 1995. – 400 с.
9. Вавилин В. А., Макарова С. И., Ляхович В, В. и др. Ассоциация полиморфных ферментов биотрансформации ксенобиотиков с предрасположенностью к бронхиальной астме у детей с наследственной отягощенностью и без таковой // Генетика. – 2002. – Т. 38. - № 4. – С. 539-545.
10. Гинтер Е.К. Популяционная генетика и медицина // Вестник РАМН. – 2001. - № 10. – С. 25-31.
11. Гланц С. Медико-биологическая статистика. – М.: Практика, 1998. — 459 с.
12. Глебович О. В. Диагностическая ценность исследования пунктата грудины при туберкулезе легких. - Ленинград., 1951. – 132 с.
13. Гончарова И. А., Фрейдин М. Б., Дунаева Л. Е., Белобородова Е. В., Белобородова Э. И., Пузырев В. П. Анализ связи полиморфизма Ile50Val гена рецептора интерлейкина-4 (IL4RA) с хроническим вирусным гепатитом // Молеклярная биология. 2005. – Т. 3. - № 3. – С. 379-384.
14. Гриппи М.А. Патофизиология легких. – М.: Восточная книжная компания, 1997. – 344 с.
15. Гусев В.А., Даниловская Е.В. Роль активных форм кислорода в патогенезе пневмокониозов // Вопр. мед. химии. – 1987. - № 5. – С. 9-15.
16. Животовский Л. А. Интеграция полигенных систем в популяциях. Проблемы анализа комплекса признаков. – М.: Наука, 1984. – 183 с.
17. Земскова З. С., Дорожкова И. Р. Скрыто протекающая туберкулезная инфекция. М.: Медицина, 1984. – 224 с.
18. Иващенко Т. Э., Сиделева О. Г., Петрова М. А. и др. Генетические факторы предрасположенности к бронхиальной астме // Генетика. – 2001. – Т. 37., № 1. – С. 107-111.
19. Ильина Н.И. Эпидемия аллергии – в чем причины? // Консилиум-медикум. – 2001. – Приложение. С. 3-5.
20. Кан Е. Л. Изменения в системе крови и их диагностическое значение // Руководство по туберкулезу органов дыхания. - 1972. — С. 116—128.
21. Крынецкий Е.Ю. Полиморфизм ферментов, участвующих в метаболизме лекарственных средств: структура генов и ферментативная активность // Молекулярная биология. – 1996. – Т.31 Выпуск 1. – 33-42.
22. Кулинский В.И. Обезвреживание ксенобиотиков // Cоросовский образовательный журнал. – 1999. - № 1. – С. 8-12.
23. Лакин Г. Ф. Биометрия: Учеб. Пособие для биол. Спец. ВУЗов – 4-е изд., перераб. И доп. М.: Высш. шк., 1990. – 352 с.
24. Лильин Е. Т., Трубников В. И., Ванюков М. М. Введение в современную фармакогенетику. – М.: Медицина, 1984. – 160 с.
25. Литвинов В. И., Чуканова В. П., Маленко А. Ф. и др. Проблемы иммуногенетики болезней легких // Сборник трудов Центр. научн-исслед. ин-та туберкулеза. – 1983. – Т. 37. – С. 16-19.
26. Литвинов В. И., Чуканова В. П., Поспелов Л. Е. и др. Роль иммуногенетических факторов при легочной патологии // Всесоюзный съезд фтизиаторов, 10-й. – Харьков, 1986. – С. 71-71.
27. Ляхович В. В., Вавилин В. А., Макарова С. И. и др. Роль ферментов биотрансформации ксенобиотиков в предрасположенности к бронхиальной астме и формировании особенностей ее клинического фенотипа // Вестник РАМН. – 2000. - № 12. – С. 36-41.
28. Ляхович В. В., Гавалов С. М., Вавилин В.А. и др. Полиморфизм генов ферментов биотрансформации ксенобиотиков и особенности бронхиальной астмы у детей // Пульмонология. 2002. – Т. 12. - № 2. – С. 31-38.
29. Ляхович В.В., Цырлов И.Б. Индукция ферментов метаболизма ксенобиотиков - Новосибирск: Наука, 1981. – 242 с.
30. Маниатис Т., Фрич Э., Сэмбук Дж. Методы генетической инженерии. Молекулярное клонирование. – М.: Мир, 1984. – 480 с.
31. Меньшиков В. В. Лабораторные методы исследования в клинике. - М.: Медицина, 1987. - 350 с.
32. Меньщикова Е.Б., Зенков Н.К. Метаболическая активность гранулоцитов при хронических неспецифических заболеваниях легких // Терапевт. арх. – 1991. - № 11. – С 85-85.
33. Милосердова А. И. Система крови при первичном туберкулезе и туберкулезном менингите у детей и ее изменение при химиотерапии: Автореф. дисс. … док-ра. мед. наук. - Кишенев, 1958. – 48 с.
34. Национальная программа «Бронхиальная астма у детей. Стратегия лечения и профилактика»: - М., 1997 г. – 93 с.
35. Огородова Л.М., Петровская Ю.А., Камалтынова Е.М. с соавт. Тяжелая бронхиальная астма у детей: факторы риска, течение // 2002. – С. 68-71.
36. Поспелов Л. Е., Серова Л. Д., Маленко А. Ф. и др. Изучение связи распределения антигенов локуса HLA-DR и туберкулеза в различных популяциях // Пробл. туб. – 1987. - № 10. – С. 54-56.
37. Проблемы наследственности при болезнях легких / Под ред. А. Г. Хоменко. – М.: Медицина, 1990. – 240 с.
38. Пузырев В. П., Фрейдин М. Б., Рудко А. А., Стрелис А. К., Колоколова О. В. Анализ взаимосвязи полиморфных маркеров генов NRAMP1 и IL12p40 и туберкулеза // Медицинская генетика. – 2002. – Т. 1. - 1. С. 44-46.
39. Пузырев В. П., Фрейдин М. Б., Огородова Л. М., Кобякова О. С. Взаимосвязь полиморфных вариантов генов интерлейкинов и их рецепторов с атопической бронхиальной астмой // Медицинская генетика. – 2002. – Т. 1. - № 2. – С. 86-92.
40. Пузырев В. П., Фрейдин М. Б., Рудко А. А., Стрелис А. К., Колоколова О. В. Полиморфизм генов-кандидатов подверженности к туберкулезу у славянского населения Сибири: пилотное исследование // Молекулярная биология. – 2002. – Т. 36. - № 5. – С. 788-791.
41. Пузырев В. П., Степанов В. А., Назаренко С. А. Геномные исследования наследственной патологии и генетическое разнообразие сибирских популяций // Молекулярная биология. 2004. – Т. 38. - № 1. – С. 129-138.
42. Пузырев В.П. Генетика мультифакториальных заболеваний: между прошлым и будущим // Медицинская генетика. – 2003. – Т. 2. № 12. – С. 498-508.
43. Пузырев В. П. Вольности генома и медицинская патогенетика // Бюл. Сиб. Медицины. – 2002. - Т. 2. – С. 16-29.
44. Пузырев В. П. Феном и гены-синтропии // Генетика человека и патология: Сб. науч. трудов / Под ред. В. П. Пузырева. – Вып. 7. – Томск: Печатная мануфактура, 2004. – 296 с.
45. Пузырев В.П., Никитин Д.Ю., Напалкова О.В. Ген NRAMP1: структура, функция и инфекционные болезни человека // Молекулярная генетика, микробиология и вирусология. – 2002. – №3. – С.34-40.
46. Пузырев В.П., Степанов В.А. Патологическая анатомия генома // Новосибирск: «Наука». – 1997. 224 с.
47. Рабухин А. Е. Туберкулез органов дыхания у взрослых. - М.: Медицина, 1976. - 328 с.
48. Радзинский А. Г. Гематологическая характеристика свежих неослажненных случаев туберкулеза легких при антибактериальной терапии // Врачебное дело. - 1961. - № 4.- С. 61-66.
49. Райс Р. Х., Гуляева Л. Ф. Биологические эффекты токсических соединений: курс лекций / Новосиб. Гос. Ун-т. – Новосибирск. – 2003. – 208 с.
50. Российская Научно-практическая программа «Бронхиальная астма у детей: диагностика, лечение и профилактика» Москва, 2004. – 46 с.
51. Рудко А.А., Ондар Э.А., Фрейдин М.Б., Пузырев В.П. Генетика подверженности к туберкулезу у тувинцев // Вестник этнической медицины. – 2004. - Т.1. - №1. – С. 17-21.
52. Сафронова О. Г., Вавилин В. А., Ляпунова А. А. Взаимосвязь между полиморфизмом гена GSTP1 и бронхиальной астмой и атопическим дерматитом // Бюл. Эксп. Биол. Мед. – 2003. – Т. 136. - № 1. – С. 73-75.
49. Сибиряк С. В. Цитокины как регуляторы цитохром Р-450 –зависимых монооксигеназ Теоретические и прикладные аспекты // Цитокины и воспаление. – 2003. - №2. – Р. 27-31.
53. Состояние противотуберкулезной помощи неселению Сибирского и Дальневосточного Федеральных округов по итогам работы в 2003 г. / Под общей редакцией Заслуженного врача Российской Федерации д.м.н., профессора В. А. Краснова. – Новосибирск. – 2004. 44 с.
54. Тиунов Л.А., Головенко Н.Я., Галкин Б.Н., Баринов В.А. Биохимические механизмы токсичности окислов азота // Успехи соврем. биологии. – 1991. – Т. 111, вып. 5. – С. 738-750.
55. Фогель Ф., Мотульски А. Генетика человека. В 3-х т./ Пер. с англ. – М.: Мир, 1990.
56. Фрейдин М.Б., Кобякова О.С., Огородова Л.М. с соавт. Наследуемость уровня общего интерлейкина-5 и полиморфизм С-703Т гена IL5 у больных бронхиальной астмой // Бюлл. Эксп. Биол. Мед. – 2000. – Т. 129 (прил. 1). – С. 50-52.
57. Фрейдин М.Б., Огородова Л.М., Пузырев В.П. Вклад полиморфизма генов интерлейкинов в изменчивость количественных факторов риска атопической бронхиальной астмы // Медицинская генетика. – 2003. - Т.2. -№ 3. – С. 130-135.
58. Хоменко А. Г., Литвинов В. И., Чуканова В. П. и др. Антигены комплекса HLA у больных туберкулезом и здоровых лиц в различных популяциях // Иммунология. – 1985. - № 1. – С. 22-24.
59. Цинзерлинг А.В., Цинзерлинг В.А. Патологическая анатомия // Учебник для педиатрических факультетов медицинских вузов. – Сотис. Санкт-Петербург. – 1996. – 369 с.
60. Чучалин А.Г. Генетические аспекты бронхиальной астмы // Пульмунология. – 1999. - № 12. – Р. 6-10.
61. Шайхаев Г.О. Туберкулез проблема не только социальная… // Природа. – 1999. - № 10. – С. 8-12.
62. Шангареева З.А., Викторова Т.В., Насыров Х.М. и др. Анализ полиморфизма генов, участвующих в метаболизме этанола, у лиц с алкогольной болезнью печени // Медицинская генетики. – 2003. - Т. 2. – № 11. – С. 485-490.
63. Шарафисламова Э.Ф., Викторова Т.В., Хуснутдинова Э.К. Полиморфизм генов глутатион S-трансфераз М1 и Р1 у больных эндометриозом из Башкортостана // Медицинская генетика. – 2003. - Т. 2. – №. 3. – С. 136-140.
64. Шмелев Н. А. Цитологический анализ крови и его значение при туберкулезе. - М., 1959. – 140 с.
65. Adjers K., Pessi T., Karjalainen J. et al. Epistatic effect of IL1A and IL4RA genes on the risk of atopy // J. Allergy Clin. Immunol. – 2004. – V. 113. - № 3. – P. 445-7.
66. Al-Arif L., Affronti L. F., Goldstein R. Predposition a la tuberculose et antigenes HLA dans une population noire de Washington // Bull. Union int. contre Tuberc. 1979. – V. 54. - № 2. – P. 151-159.
67. Alexandrie A.K., Ingelman-sundberg M., Seidegaard J. et al Genetic susceptibility to lung cancer: a study of host factors in relation to age of onset and histological cancer types // Carcinogenesis (Lond.). – 1994. – V. 15. – P. 1785-1790.
68. Anderson G. G., Cookson W. O. C. M. Recent advances in the genetics of allergy and asthma // Mol. Med. Today. - 1999. – V. 5. – P. 264-273.
69. Anttila S., Luostarinen L., Hirvonen A. et al. Pulmonary expression of glutathione S-transferase M3 in lung cancer patients: assotiation with GSTM1 polymorphism, smoking, and asbestos exposure // Cancer Res. – 1995. - V. 55.– P. 3305-3309.
70. Arai K.I., Lee F., Miyajima A. et al. Cytokines co-ordinators of immune and inflammatory responses // Ann. Rev. Biochem. – 1990. – V. 59. – P. 783-802.
71. Awasthi S. S., Srivastava F. K., Ahmad F. et al. Interaction of glutathione S-transferase-pi with ethacrynic acid and its glutathionic conjugate // Biochem. Biophys. Acta. – 1993. – V. 1164. – P. 173-178.
72. Baldini M., Lohman I.C., Halonen M et al. A Polymorphism in the 5’ flanking region of the CD14 levels and with total serum immunoglobulin E // Am. J. Respir. Cell Mol. Biol. – 1999. – V. 20. – P. 976 -983.
73. Bartsch H., Nair U., Risch A. et al. Genetic polymorphism of CYP genes, alone or in combination, as a risk modifier of tobacco-related cancers // Cancer Epidemiology, Biomarkers and Prevention. – 2000. – V. 9. – P. 3-28.
74. Beckett G.J., Hayes J.D. Glutathione S-transferases: biomedical applications // Adv. Clin. Chem. – 1993. – V. 30. – P. 281-380.
75. Bellamy R. Identifyng genetic susceptibility factors for tuberculosis in African: a combined approach using a candidate gene study and a genome-wide screen // Clinical Science. – 2000. – V. 98. – P. 245-250.
76. Bellamy R., Ruwende C., Corra T. et al. Variation in the NRAMP1 gene and susceptibility to tuberculosis in West Africans // The New England Journal of Medicine. 1998. – V. 338. - № 10. – P. 640-644.
77. Bertz R. J., Granneman G. R. Use of in vitro and in vivo date to estimate the likelihood of metabolic pharmacokinetic interactions // Clin Pharmacokinet. 1997. – V. 32. – P. 210-258.
78. Board P.G., Webb G.C., Coggan M. Isolation of cDNA clone and localization of the human glutathione S-transferase 3 on chromosome bands 11q13 and 12q13-14 // Ann. Hum. Genet. – 1989. – V. 53. – P. 205-213.
79. Bornman L., Campbell S. J., Fielding K. et al. Vitamin D receptor polymorphisms and susceptibility to tuberculosis in West Africa: a case-control and family study // J. Jnfect. Dis. – 2004. - V. 190. - № 9. – P. 1631-1641.
80. Brasch-Andersen C, Christiansen L, Tan Q. Possible gene dosage effect of glutathione-S-transferases on atopic asthma: using real-time PCR for quantification of GSTM1 and GSTT1 gene copy numbers // Hum Mutat. – 2004. – V.24. - № 3. – Р. 208-214.
81. Brockmoller J., Cascorbi I., Kerb R. Combined analysis of inherited polymorphisms in arylamine N-acetyltransferase 2, glutathione S-transferase M1 and T1, microsomal epoxide hydrolase, and cytochrome P450 enzymes as modulators of bladder cancer risk // Cancer Res. – 1996. – V. 56. – P. 3915-3925.
82. Burchard E. G., Silverman E. K., Rosenwasser L. J. et al. Assotiation between a sequence variant in the IL4 promoter and FEV(1) in asthma // Am. J. Respir. Crit. Care Med. – 1999. - № 160. – P. 919-922.
83. Cannone-Hergaux F., Gruendheid S. et al The NRAMP1 protein and its role resistence to infection and makrophage funktion // Proc. Amer. Physicians. – 1998. – V. 111. - № 4. – P. 283-289.
84. Carroll W.D., Lenney W., Child F. et al. Maternal glutathione S-transferase GSTP1 genotype is a specific predictor of phenotype in children with asthma // Pediatr Allergy Immunol.- 2005. - V.1. - № 16. – P. 32-39.
85. Carter C.O. Polygenic inheritance in man // Br. Med. Bull. – 1996. – V. 25. – P. 52-57.
86. Cervino A. C. L., Lakiss S., Sow O. et al. Allelic assotiation between the NRAMP1 gene and susceptibility to tuberculosis in Guinea-Conakry // Ann. Hum. Genet. 2000. – V. 64. – P. 507-512.
87. Chen H., Sandler D.P., Taylor J.A. et al. Increased risk for myelodysplastic syndromes in individuals with glutathione transferase theta 1 (GSTT1) gene defect // The lancet. – V. 347. – 1996. – P. 295-297.
88. Chung K.F., Barnes P.J. Cytokines in asthma // Thorax. – 1999. – V. 54. – P. 825-857.
89. Cristina E. Mapp MD, Anthony A.et al. Glutathione S-transferase GSTP1 is a susceptibility gene for occupational asthma induced by isocyanates // Environmental and Occupational Disorders. – 2002.
90. Crump C., Chen C., Appelbaum F.R. et al. Glutathione S-transferase theta 1 gene deletion and risk of acute myeloid leukemia // Cancer Epidemiology, Biomarkers & Prevention. – V. 9. 2000. – P. 457-460.
91. Cytochrome P450 [Электронный ресурс]. – Режим доступа: http://drnelson.utmem.edu/Cytochrome P450.html/
92. Daniels S.E., Bhattacharrya B., James A. et al. A genome-wide search for quantitative trait loci underlying asthma. // Nature. – 1996. – V. 383. – P. 247-250.
93. De Long J.L., Chang T.M., Whang-Peng J. et al. The human liver glutathione S-transferase gene superfamily: expression and chromosome mapping of an Hb subunit cDNA // Nucleic. Acid Res. – 1988. – V. 16. – P.8541-8554.
94. De Morais S. M. F, Wilkinson G. R., Blaisdell J. et al. The major genetic defect responsible for the polymorphism of S-mephenytoin metabolism in human // J. Biol. Chem.-1994.- V. 269.- №22. - P. 15419-15422.
95. Delfino R.J., Sinha R., Smith S. et al., Breast cancer, heterocyclic aromatic amines from meat and N-acetyltransferase 2 genotype // Carcinogenesis. – 2000. – V. 21. – P. 607-615.
96. Denison M.S. Whitlock J.P.Jr. Xenobiotic-inducible transcription of cytochrome P450 genes // J. Biol. Chem. - 1995. – V. 270. – P. 18175-18178.
97. Dickinson D.S., Bailey W.C., Hirschowihz B.I. et al. Risk factors for isiniazid induced liver dysfunction // J. Clin Gastroenterol. – 1981. – V. 3. – P. 271-279.
98. Directory of P450-containing systems [Электронный ресурс]. – Режим доступа: http://www.icgeb.trieste.it/~p450srv/
99. Dirksen U., Moghadam K. A., Mambetova C. et al. Glutathione S-transferase theta 1 gene (GSTT1) null genotype is associated with an increased risk for acquired aplastic anemia in children // Pediatric Research. – 2004. – V. 55. – P. 466-471.
100. Dizier M. H., Sandford A., Walley A. et al. Indication of linkage of serum IgE levels to the interleukin-4 gene and exclusion of the contribution of the (-590 C to T) interleukin-4 promoter polymorphism to IgE variation // Genet. Epidemiol. – 1999. - № 16. P. 84-94.
101. Drysdale C. M., McGraw D. W., Stack C. B. et al. Complex promoter and coding region beta 2-adrenergic receptor haplotypes alter receptor expression and predict in vivo responsiveness // Proc. Nat. Acad. Sci. USA. – 2000. - № 97. – P. 10483-10488.
102. Duffy D. L., Martin N. G., Battistutta D. et al. Genetics of asthma and hay fever in Australian Twins // Am. Rev. Respir. Dis. – 1990. – V. 142. - № 6 (Pt. 1). – P. 1351-1358.
103. Eaton D.L., Bammler T.K. Concise review of the glutathione S-transferases and their significance to toxicology // Toxicol. Sci. – 1999. – V. 49. – P. 156-164.
104. Eder W, Klimecki W, Yu L et al. Toll-like receptor 2 as a major gene for asthma in children of European farmers // J. Allergy Clin. Immunol. - 2004. – V. 113. - 3. – P. - 482-488.
105. Edfors-Lubs M. L. Allergy in 7000 twin pairs // Acta Allergol. – 1971. – V. 26. - № 4. – P. 249-285.
106. Engel G.,Hofman U., Heidemann H. et al. Antipyrine as a probe for human oxidative drug metabolism: identification of the cytochrome P450 enzymes catalyzing 4-hydroxyantipyrine, 3- hydroxymethylantipyrine, and norantipyrine formation // Clin. Pharmacol. Ther. – 1996. – V. 59. - № 6. – P. 613-623.
107. Evans W. E., Relling M. V. Pharmacogenomics: translating functional genomics into rational theurapeutics // Science. – 1999. – V. 286. – P. 487-491.
108. Fageras Bottcher M., Hmani-Aifa M., Lindstrom A. et al. A TLR4 polymorphism is associated with asthma and reduced lipopolysaccharide-induced interleukin-12(p70) responses in Swedish children // J. Allergy Clin. Immunol. – 2004. – V. 114. - № 3. – P. 561-567.
109. Farker K., Lehmann M. H., Oelschlagel B. et al. Impact of CYP2E1 genotype in renal cell and urothelial cancr patients // Exp. Toxicol. Pathol. – 1998. - V. 50. – P. 425-431.
110. Frayer A.A., Hume R., Strange R.C. The development of glutathione S-transferase and glutathione peroxidase activities in human lung // Biochim. Biophys. Acta. – 1986. - № 883. P. 448-453.
111. Freidin M.B., Kobyakova O.S., Ogorodova L.M. et al. Association of polymorphisms in the human IL4 and IL5 genes with atopic bronchial asthma and severity of the disease // Comp. Funct. Genom. – 2003. - № 4. – P. 346-350.
112. Freimer N., Sabatti C. The human phenome project // Nat. Genet. – 2003. – V. 34. - № 1. – P. 15-21.
113. Frodsham A. J., Hill A. S. Genetics of infections diseases // Hum. Mol. Genet. – 2004. – V. 13. – Review Issue 2. – P. 187-194.
114. Fryer A. A., Bianco A., Hepple M. et al. Polymorphism at the glutathione S-transferase GSTP1 locus. A new marker for bronchial hyperresponsiveness and asthma // Am. J. Respir. Crit. Care Med. – 2000. – V. 161. – P. 1437-1442.
115. Fukai H., Ogasawara Y., Migita O. et al. Association between a polymorphism in cysteinyl leukotriene receptor 2 on chromosome 13q14 and atopic asthma // Pharmacogenetics. – 2004. – V. 14. - №10. – P. - 683-90.
116. Gao P. S., Mao X. Q., Baldini M. et al. Serum total IgE levels and CD14 on chromosome 5q31 // Clin. Genetic. – 1999. – V. 56. – P. 164-165.
117. Gao P.-S., Fujishima S., Mao X.-Q., et al. Genetic variants of NRAMP1 and active tuberculosis in Japanese populations // Clin. Genet. – 2000. – V. 58. – P. 74-76.
118. Gawronska-Sklarz B., Pawlik A., Czaja-Bulsa G. et al. Genotype of N-acetyltransferase 2 (NAT2) polymorphism in children with immunoglobulin E-mediated food allergy // Clin. Pharmacol. Ther. – 2001. – V. 69. - № 5. – P. 372-378.
119. Gilliand F. D., Li Y.-F., Dubeau L. et al. Effects of glutathione S-transferase M1, maternal smoking during pregnancy, and environmental tobacco smoke on asthma and wheezing in children // Am. J. Respir. Crit Care Med. – 2002. – V. 166. – P. 457-463.
120. Gonzalez F.J. Molecular biology and regulation of phase I enzymes // Abstr. of 5th European ISSX Meeting, Tours. September, 26-29. – 1993. – V. 3. – P. 139.
121. Graves P. E., Kabesch M., Halonen M. et al. A cluster of seven tightly linked polymorphisms in the IL-13 gene associated with total serum IgE levels in three populations of white children // J. Allergy Clin. Immunol. – 2000. - № 105. – P. 506-513.
122. Greenwod C.M.T., Fujiwara M.T. Linkage of tuberculosis to chromosome 2q35 loci, including NRAMP1, in a large aboriginal Canadian family // Am. J. Hum. Genet. – 2000. – V. 67. – P. 405-416.
123. Guengerich F.P. Cytochrome P450s, drugs and diseases // Molecular Interventions. – 2003. – V. 3. № 4. – P. 8-18.
124. Guengerich F.P. Role of cytochrome P450 enzymes in chemical carcinogenesis and cancer chemotherapy // Cancer Res. – 1988. – V. 48. – P. 2946-2954.
125. Haehner B.D, Gorski J.C., Vandenbranden M. et al. Bimodal distribution of renal cytochrome P450 3A activity in humans // Mol. Pharmacol. – 1996. – V. 50. – P. 52-59.
126. Hall I. P., Wheatley A., Christine G. et al. Assotiation of CCR5 delta32 with reduced risk of asthma // Lancet. – 1999. - № 354. – P. 1264-1265.
127. Hallier E., Langhof T., Dannappel D. et al. Polymorphism of glutathione conjugation of methyl bromide, ethylene oxide and dichloromethane in human blood: influence on the induction of sister chromatid exchanges (SCE) in lymphocytes // Arch. Toxicol. – 1993. V. 67. - № 3. – P. 173-178.
128. Hasegawa K., Tamari M., Shao C. et al. Variations in the C3, C3a receptor, and C5 genes affect susceptibility to bronchial asthma // Hum Genet. – 2004. – V. 115. - № 4. P. - 295-301.
129. Hayashi S., Watanabe J., Kawajiri K. Genetic polymorphisms in the 5’-flanking region change transcriptional regulation of the human cytochrome P450IIE1 gene // J. Biochem. – 1991. – V. 110. - P. 559-565.
130. Hayes J. D., McLellan L. I. Glutathione and glutathione-dependent enzymes represent a co-ordinately regulated defence against oxidative stress // Free Radic. Res. – 1999. – V. 31. P. 273-300.
131. Hayes J. D., Strange R. C. Glutathione S-transferase polymorphisms and their biological consequences // Pharmacology. – 2000. – V. 61. – P. 154-166.
132. Heinzmann A., Mao X. Q., Akaiwa M. et al. Genetic variants of IL-13 signalling and human asthma and atopy // Hum. Mol. Genet. – 2000. - № 9. – P. 549-559.
133. Heinzmann A., Mao X. Q., Akaiwa M. et al. Genetic variants of IL-13 signalling and human asthma and atopy // Hum. Mol. Genet. – 2000. - № 9. – P. 549-559.
134. Heinzmann A., Plesnar C., Kuehr J. et al. Common polymorphisms in the CTLA-4 and CD28 genes at 2q33 are not associated with asthma or atopy // Eur J Immunogenet. – 2000. – V. 27. – P. 57-61.
135. Hershey G. K., Friedrich M. F., Esswein L. A. et al. The association of atopy with a gain-of-function mutation in the alpha subunit of the interleukin-4 receptor // N. Engl. J. Med. 1997. - № 337. – P. 1720-1725.
136. Hill A. V. S. The immunogenetics of human infection diseases // Ann. Rev. Immunol. – 1998. - 16. – P. 593-617.
137. Hill M. R. Cookson W. O. A new variant of the beta subunit of the high-affinity receptor for immunoglobulin E (Fc epsilon RI-beta E237G): associations with measures of atopy and bronchial hyper-responsiveness // Hum. Mol. Genet. 1996. - № 5. – P. 959-962.
138. Hill M. R., James A. L., Faux J. A. et al. Fc epsilon RI-beta polymorhism and risk of atopy in general population sample // BMJ. – 1995. - № 311. – P. 776-779.
139. Hirota T., Obara K., Matsuda A. et al. Association between genetic variation in the gene for death-associated protein-3 (DAP3) and adult asthma // J. Hum. Genet. 2004. – V. 49. - № 7. – P. - 370-375.
140. Hizawa N., Freidhoff L. R., Chiu Y. F. et al. Genetic regulation of Dermatophagoides pteronyssinus-specific IgE responsiveness: a genome-wide multipoint linkage analisis in families recruited through 2 asthmatic sibs. The Collaborative Study on the Genetics of Asthma (CSGA) // J. Allergy Clin Immunol. – 1998. - V. 102. – P. 436-442.
141. Hoffjan S, Ostrovnaja I, Nicolae D. et al. Genetic variation in immunoregulatory pathways and atopic phenotypes in infancy // J. Allergy Clin. Immunol. – 2004. V. 113. - № 3. – P. - 511-518.
142. Holgate S.T., Cherch M.K., Howarth P.H. et al., Genetic and environmental influences on airway inflammation in asthma // Int. Arch. Allergy Immunol. - 1995. – V. 107. – P. 29-33.
143. Holla L.I., Schuller M., Buckova D. et al. Neuronal nitric oxide synthase gene polymorphism and IgE-mediated allergy in the Central European population // Allergy. – 2004. – V. 59. - № 5. – P. - 548-552.
144. Honkakoski P., Negishi M. Regulation of cytochrome P450 (CYP) genes by nuclear receptors // Biochem. J. 2000. – V. 347. – P. 321-337.
145. Hu Y., Oscarson M., Johanson I. et al. Genetic polymorphism of human СYP2E1: a characterization of two variant alleles // Mol. Pharmacol. 1997. – V. 51. – P. 370-376.
146. Huang S. K., Marsh D. G. Genetics of allergy // Ann. Allergy. – 1993. – V. 70. – P. 347-359.
147. Huang Y. S., Chern H. D., Su W. J. et al. Polymorphism of the N-acetyltransferase 2 gene as a susceptibility risk factor for antituberculosis drug-induced hepatits // Hepatology. – 2002. – V. 35. - № 4. – P. 883-889.
148. Huang Y.-S., Chern H.-D., Su W.-J. et al. Cytochrome P450 2E1 genotype and the susceptibility to antituberculosis drug-induced hepatitis // Hepatology. – 2003. – V. 37. - № 4. P. 924-930.
149. Hytonen A. M, Lowhagen O., Arvidsson M. et al. Haplotypes of the interleukin-4 receptor alpha chain gene associate with susceptibility to and severity of atopic asthma // Clin Exp Allergy. – 2004. – V. 34. - № 10. - P. - 1570-1575.
150. Ibenau G.C., Blaisdell J., Chanayem B.I et al. An additional defective allele, CYP2C19*5, contributes to the S-maphenytoin poor metabolizer phenotype in Caucasians // Pharmacogenetics. 1998. – V. 8. - № 2. – P. 129-135.
151. Ibenau G.C., Blaisdell J., Ferguson R. J. et al. A novel transversion in the intron 5 donor splice junction of CYP2C19 and a sequence polymorphism in exon 3 contribute to the poor metabolizer phenotype for the anticonvulsant drug S-mephenytoin // The J. Pharmacol. Exp. Ther. – 1999. – V. 290. - № 2. – P. 635-640.
152. Ingelman-Sundberg M. Human drug metabolizing cytochrome P450 enzymes: properties and polymorphisms // Arch Pharmacol. – 2004. – V. 369. – P. 89-104.
153. Ingelman-Sundberg M. Pharmacogenetics of cytochrome P450 and its applications in drug therapy: the past, present and future // Trends Pharmacol Sci. – 2004. – V. 25. - №. 4. – P. 193-200.
154. Ingelman-Sundberg M., Oscarson M., Persson I. et al. Genetic polymorphism of human drug metabolizing enzymes. Recent aspects on polymorphic forms of cytochromes P450 // European Commission. European cooperation in the field of scientific and technical research. COST B1 Conference on variability and specificity in drug metabolism. Edited by Alvan G. Brussels. – 1995. – P. 93-110.
155. Ioannides C., Lewis D. F. Cytochromes P450 in the bioactivation of chemicals // Curr. Top. Med. Chem. 2004. – V. 4. - № 16. – P. 1767-1788.
156. Ishii T., Matsuse T., Teramoto S et al. Glutathione S-transferase P1 (GSTP1) polymorphism in patients with chronic obstructive pulmonary disease // Thorax. – 1999. – V.54. – P. 693-696.
157. Jang N., Stewart G., Jones G. et al. Polymorphisms within the PHF11 gene at chromosome 13q14 are associated with childhood atopic dermatitis // Genes Immun. – 2005. – V. 6. - № 3. – P. 262-264.
158. Ji X., Johnson W.W., Sesay M.A. et al. Structure and function of the xenobiotic substrate binding site of a glutathione S-transferase as revealed by X-ray crystallographic analysis of product complexes with the diasteriomers of 9-(S-glutathionyl)-10-hydroxy-9,10-dihydrophenantrene // Biochemistry. – 1994. V. 33. – P. 1043-1052.
159. Jiang Z. F., Aw J. B., Sun Y. P. et al. Assotiation of HLA-Bw35 with tuberculosis in the Chinese // Tiss. Antigens. – 1983. – V. 22. - № 1. - P. 86-88.
160. Jourenkowa-Mironova N., Wikman H., Bouchardy C. et al. Role of glutathione S-transferase GSTM1, GSTM3, GSTP1 and GSTT1 genotypes in modulating susceptibility to smoking-related lung cancer // Pharmacogenetics. – 1998. – V. 8. – P. 495-502.
161. Kabesch M., Carr D., Weiland S. K. et al. Association between polymorphisms in serine protease inhibitor, kazal type 5 and asthma phenotypes in a large German population sample // Clin. Exp. Allergy. – 2004. – V. 34. - № 3. – P. 340-345.
162. Katoh T., Kaneko S., Jakasawa S. et al. Human glutathione S-transferase P1 polymorphism and susceptibility to smoking related epithelial cancer; oral, lung, gastric, colorectal and urothelial cancer // Pharmacogenetics. – 1999. -V. 9. – P. 165-169.
163. Kawashima T., Noguchi E., Arinami T. et al. Linkage and association of an interleukin 4 gene polymorphism with atopic dermatitis in Japanese families // J. Med. Genet. – 1998. - № 35. P. 502-504.
164. Kedda M. A, Shi J., Duffy D. et al. Characterization of two polymorphisms in the leukotriene C4 synthase gene in an Australian population of subjects with mild, moderate, and severe asthma // J. Allergy Clin. Immunol. – 2004. – V. 113. - 5. – P. - 889-95.
165. Kharasch E. D., Thummel K. E., Mhzyre J. et al. Single-dose disulfiram inhibition of chlorzoxazone metabolism // Clin Pharmacol Ther. – 1993. – V. 53. – P. 643-650.
166. Kihara M., Noda K. Lung cancer risk of the GSTM1 null genotype is enhanced in the presence of the GSTP1 mutated genotype in male Japanese smokers // Cancer Lett. – 1999. – V. 137. P. 53-60.
167. Kolble K. Regional mapping of short tandem repeats on human chromosome 10: cytochrome P450 gene CYP2E1, D10S196, D10S220, and D10S225 // Genomics. -1993. - V. 18. P. 702-704.
168. Koppelman G. H., Stine O. C., Xu J. et al. Genome-wide search for atopy susceptibility genes in Dutch families with asthma // J. Allergy Clin. Immunol. - 2002. - V. 109. - P. 498-506.
169. Kruse S., Japha T., Tender M. et al. The polymorphisms S503P and Q576R in the interleukin-4 receptor alpha gene are associated with atopy and influence the signal transduction // Immunology. – 1999. - № 96. – P. 365-371.
170. Kruse S., Mao X. Q., Heinzmann A. at al. The Ile198Thr and Ala379Val variants of plasmatic PAF-acethylhydrolase impair catalytical activities and are associated with atopy and asthma // Am. J. Hum. Genet. – 2000. – V. 66. – P. 1522-1530.
171. Lahiri D.K., Bye S., Nunberg J.I., et al. Anon-organic and non-enzymatic eztraction method gives higher yields of genomic DNA from whole-blood samples than do nine other methods used // J. of Biochemical and Biophysical Methods. – 1992. – V. 25. – P. 193-205.
172. Laing I. A., Goldblatt J., Eber E. et al. A polymorphism of the CC16 gene is associated with an increased risk of asthma // J. Med. Genet. – 1998. – V. 35. P. 463-467.
173. Laitinen T., Rasanen M., Kaprio J. et al. Importance of Genetic factors in adolescent asthma. A population-based twin-family study // Am. J. Respir. Crit. Care Med. – 1998. V. 157. – P. 1073-1078.
174. LeSouef P. Genetics of asthma: What do we need to know? // Pediatr. Pulmonol. – 1997. – Suppl. 15. P. 3-8;
175. Lewis D. F., Lake B. G., Dickins M. Substrates of human cytochromes P450 from families CYP1 and CYP2: analysis of enzyme selectiveity and metabolism. – 2004. – V. 20. - № 3. – P. 111-142.
176. Lichtenstein P., Svartengren M. Genes, environments, and sex: factors of importance in atopic diseases in 7-9-Year-old Swedish twins // Allergy. – 1997. – V. 52. - № 11. P. 1079-86.
177. Lin D.-X., Tang Y.-M., Peng Q. et al. Susceptibility to esophageal cancer and genetic polymorphisms in glutathione S-transferases T1, P1 and M1 and cytochrome P450 2E1 // Cancer Epidemiol. Biomarkers Prev. – 1998. – V. 7. – P. 1013-1018.
178. Liu X., Nickel R., Beyer K. et al. An IL-13 coding region variant is associated with a high total serum IgE level and atopic dermatitis in the German multicenter atopy study (MAS-90). // J. Allergy Clin. Immunol. – 2000. - № 106. – P. 167-170.
179. Lomas D. A., Silverman E. K. The genetics of chronic obstructive pulmonary disease // Respir. Res. 2001. – V. 2. - № 1. – P. 20-26.
180. Luszawaka-Kutrzela T. NAT2 genotype in children with bronchial asthma and other atopic diseases // Ann. Acad. Med. Stein. – 1999. – V. 45. – P. 109-21.
181. Mace K., Bowman E.D., Vautravers P. et al. Characterisation of human xenobiotic-metabolizing enzymes expression in human bronchial mucosa and peripheral lung tissues // Europ. J. Cancer. – 1998. – V. 34. – P. 914-920.
182. MacLeod S., Sinha R., Kadlubar F.F. et al. Polymorphisms of CYP1A1 and GSTM1 influence the in vivo function of CYP1A2 // Mutat. Res. – 1997. - V. 376. – P. 135-142.
183. Mao X. Q., Shiracawa T., Yoschikawa K. et al. Association between genetic variants of mast-cell chymase and eczema // Lancet. – 1996. - № 348. – P. 581-583.
184. Matsushita I., Hasegawa K., Nakata K. et al. Genetics Variants of Human b-Defensin-1 and Chronic Obstructive Pulmonary Disease // Biochemical and Biophysical Research Communications. – 2002. – V. 291. – P. 17-22.
185. Melen E., Bruce S., Doekes G. et al. Haplotypes of G-protein-coupled Receptor 154 are Associated with Childhood Allergy and Asthma // Am. J. Respir. Crit. Care Med. – 2005. – V. 121. – P. 1089-1095.
186. Meyer C.G., May J., Stark K. Human Leukocyte antigens in tuberculosis and leprosy // Trends Microbiol. 1998. - V. 6. - № 4. – P. 148-154.
187. Miller D.P., Liu G., De Vivo I. et al. Combinations of the variant genotypes of GSTP1, GSTM1 and p53 are associated with an increased lung cancer risk // Cancer Research. – 2002. - V. 62. – P. 2819-2823.
188. Mitsuyasu H., Izuhara K., Mao X.Q. et al. Ile50Val variant of IL4R alpha upregulates IgE synthesis and associates with atopic asthma // Nat. Genet. – 1998. - № 19. – P. 119-120.
189. Moffatt M. F., Cookson W. O. Tumor necrosis factor haplotypes and asthma // Human Molecular Genet. – 1997. – V. 6. – P. 551-554.
190. Moffatt M. F., Schou C., Faux J. A. et al. Germline TCR-A restriction of immunoglobulin E responses to allergen // Immunogenetics. – 1997. - № 46. – P. 226-230.
191. Morgenstern R., DePierre J. W. Microsomal glutathione S-transferase // Rev. Biochem. Toxicol. - 1985. V. 7. – P. 67-103.
192. Morita S., Yano M., Shiozaki et al. CYP1A1, CYP2E1 and GSTM1 polymorphisms are not associated with susceptibility to squamouscell carcinoma of the esophagus // Int. J. Cancer. 1997. – V. 71. – P. 192-195.
193. Munaka M., Kohshi K., Kawamoto T et al. Genetic polymorphisms of tobacco- and alcohol-related metabolizing enzymes and the risk of hapatocellular carcinoma // Journal of cancer research and clinical oncology. – 2003. - V. 129. – №. 6. - P. 355-360.
194. Mutmansky J.M. The war on black lung // Earth and Miner. Sci. – 1990. – V. 59. – P. 6-10.
195. Nanavaty U., Goldstein A.D., Levine S.J. Polymorphisms in candidate asthma genes // Am. J. Med. Sci. 2001. – V. 321. – P. 11-16.
196. Nebert D. W., Jorge-Nebert L., Vesell E. S. Pharmacogenomics and “Individualized drug therapy” // Am. J. Pharmacogenomics. – 2003. – V. 3. - № 6. – P. 361-370.
197. Nebert D.W. Polymorphisms in drug-metabolizing enzymes: what is their clinical relevance and why do they exist? // Am. J. Hum. Genet. – 1997. - № 60. – P. 265-271.
198. Nelson D. R., Koymans L., Kamataki T. et al. P450 superfamily: update on new sequences, gene mapping, accession numbers and nomenclature // Pharmacogenetics. – 1996. – V. 6. – P. 1-42.
199. Nickel R. G., Casolaro V., Wahn U. et al. Atopic dermatitis is associated with a functional mutation in the promoter of the C-C chemokine RANTES // J. Immunol. – 2000. - 164. – P. 1612-1616.
200. Nicolae D., Cox N.J., Lester L. et al. A fine mapping and positional candidate studies identify HLA-G as an asthma susceptibility gene on chromosome 6p21 // Am. J. Hum. Genet. – 2005. – V. 76. - № 2. – P. - 349-57.
201. Nieminen M. M., Kaprio J., Koskenvuo M. A population-based study of bronchial asthma in adult twin pairs // Chest. – 1991. – V. 100. – P. 70-75.
202. Noguchi E., Shibasaki M., Arinami T. et al. Assotiation of asthma and the interleukin-4 promoter gene in Japanese // Clin Exp Allergy. – 1998. – 28. – P. 449-453.
203. North R .J., Medina E. How important is Nramp1 in tuberculosis? // Trends in Microbiology. – 1998. – V. 6. - №11. – P. 441-443.
204. Obase Y., Shimoda T., Kawano T. et al. Polymorphisms in the CYP1A2 gene and theophylline metabolism in patients with asthma // Clin Pharmacol Ther. – 2003. – V. 73. - № 5. – P. 468-474.
205. Ober C., Cox N. J., Abney M. et al. Genome-wide search for asthma susceptibility loci in a founder population. The Collaborative Study on the Genetics of Asthma // Hum. Mol. Genet. – 1998. – V. 7. - P1393-1398.
206. Ober S., Leavitt S. A., Tsaienko A. et al., Variation in the interleukin 4-receptor alpha gene confers susceptibility to asthma and atopy in ethnically diverse populations // Am. J. Hum. Genet. – 2000. - № 66. – P. 517-526.
207. Organov R.G., Maslennikova G. Ya. // Eur. Respir. J. – 1999. – V. 13. - № 2. – P. 287- 289.
208. Paigen K.and Eppig J. T. A mouse phenome project / Mamm. Genome. – 2000. – V. 11. - № 9. – P. 715-717.
209. Park J.Y., Schantz S.P., Stern J.C. et al., Assotiation between glutathione S-transferase pi genetic polymorphism and oral cancer risk // Pharmacogenerics. – 2000. - V. 10. - № 4. P. 374.
210. Pearce N. What does the odds ratio estimate in a case-control study? // Int. J. Epidemiol. – 1993. – V. 26 № 6. – P. 1189-1192.
211. Pelkonen O., Raunio H. Metabolic activation of toxins: tissue-specific expression and metabolism in target organs // Environ. Health Perspect. – 1997. – V. 105. – Suppl. 4. – P. 767-774.
212. Pemble S., Schroeder K.R., Spencer S.R. et al. Human glutathione S-transferase theta (GSTT1): cDNA cloning and the characterization of a genetic polymorphism // Biochem J. – 1994. – V. 300. – P. 271-276.
213. Pillai S. G, Cousens D. J, Barnes A. A. A coding polymorphism in the CYSLT2 receptor with reduced affinity to LTD4 is associated with asthma // Pharmacogenetics. 2004. – V. 14. - № 9. – P. - 627-633.
214. Poland A., Glover E., Robinson J. R. et al. Genetic expression of aryl hydrocarbon hydroxylase activity, induction of monooxygenase activities and cytochrome P450 formation by 2, 3, 7, 8-tetrachlorodibenzo-p-dioxin in mice genetically “non-resposive to others aromatic hydrocarbons // J. Biol. Chem. – 1973. – V. 249. – P. 5599-5606.
215. Prandota J. Important role of prodromal viral infection responsible for inhibition of xenobiotic metabolizing enzymes in the pathomechanism of idiopathic Reye’s syndrome, Stevens-Johanson syndrome, autoimmune hepatits and hepatotoxicity of therapeutic doses of acetaminophen used in genetically predposed persos // Am. J. Ther. – 2002. – V. 9. – P. 149-156.
216. Pykalainen M, Kinos R, Valkonen S. et al. Association analysis of common variants of STAT6, GATA3, and STAT4 to asthma and high serum IgE phenotypes // J. Allergy Clin. Immunol. – 2005. – V. 115. - № 1. – P. - 80-87.
217. Quanjer P.H., Tammeling G.L., Cotes J.E. et al. Lung volumes and ventilatory flows // Eur. Respir. J. 1993. – V. 6. – P. 4-40.
218. Ramsey C.D., Lazarus R., Camargo C.A. et al. Polymorphisms in the interleukin 17F gene (IL17F) and asthma // Genes Immun. – 2005. – V. 6. - № 3. – P. 236-241.
219. Raunio H., Husgafvel-Pursianen, Anttila S. et al. Diagnosis of polymorphisms in carcinogen-activating and inactivating enzymes and cancer susceptibility – a review // Gene. – 1995. – V. 159. – P. 113-121.
220. Ravindranath V. Metabolism of xenobiotics in the central nervous system: implications and challenges // Biochem. Pharmacol. – 1998. – V. 56. – P. 547-551.
221. Reihsaus E., Innis M., Macintyre N. et al. Mutations in the gene encoding for the beta 2-adrenergic receptor in normal and asthmatic subjects // Am. J. Respir. Cell Mol. Biol. – 1993. – V. 8. – P. 334 -339.
222. Renton K.W. Cytochrome P450 regulation and drug biotransformation during inflammation and infection // Curr Drug Metab. – 2004. - V. 5. № 3. – P. 235-243.
223. Richardson T.H., Jung F., Griffin K.J. et al. A universal approach to the expression of human and rabbit cytochrome P450s of the 2C subfamily in Escherichia coli // Arch. Biochem. Biophys. – 1995. – V. 323. - № 1. – P. 87-96.
224. Richter-Hintz D., Their R., Steinwachs S. et al. Allelic variants of drug metabolizing enzymes as risk factors in psoriasis // Journal of Investigative Dermatology. – 2003. - V. 120. P. 765-770.
225. Rifkind A. B., Lee C., Chang T. K. and Waxman D. J. Arachidonic acid metabolism by human cytochrome P450s 2C8, 2C9, 2E1, and 1A2: regioselective oxygenation and evidence for a role for CYP2C enzymes in arachidonic acid epoxygenation in human liver microsomes // Arch Biochem Biophys. –1995. – V. 320. - № 3. – P. 380-389.
226. Rollinson S., Levene A. P., Mensah F. K., Roddam P. L. et al. Gastric marginal zone lymphoma is associated with polymorphisms in genes involved in inflammatory response and antioxidative capacity // Blood. – 2003. - V. 102. - № 3. – P. 1007-1011.
227. Romkes M., Faletto M.B., Blaisdell I.A. et al. Cloning and expression of the complementary DNAs for multiple members of the human cytochrome P450IIC subfamily // Biochemistry. 1991. – V. 30. № 13. – P. 3247-55.
228. Rosenwasser L. J., Klemm D. J., Dresback J. K. et al. Promoter polymorphisms in the chromosome 5 gene cluster in asthma and atopy // Clin. Exp. Allergy. – 1995. - № 25 (suppl 2). P. 74-78.
229. Roy B., Chowdhury A., Kundu S. et al. Increased risk of antituberculosis drug-induced hepatotoxicity in individuals with glutathione S-transferase M1 “null” mutation // Journal of Gastroenterology and Hepatology. – 2001. – V. 16. – P. 1033-1037.
230. Ryu S., Park Y.-K., Bai G. H. et al. 3’ UTR polymorphisms in the NRAMP1 gene are associated with susceptibility to tuberculosis in Koreans // Int. J. Tuberc. Lung. Dis. – 2000. V. 4. - № 6. – P. 577-580.
231. Saito K. Shinoharaet A., Kamataki T. et al. N-hydroxylanine O-acetyltransferase in hamster liver: identity with arylhydroxamic acid N,O-acetyltransferase and arylamine N- acetyltransferase // J. Biochem. – 1986. – V. 99. - № 6. – P. 1689-1697.
232. Salama S. A., Sierra-Torres C. H., Oh H-Y. et al. A multiplex-PCR/RFLP procedure for simultaneous CYP2E1, mEH and GSTM1 genotyping // Cancer Lett. – 1999. – V. 143. P. 51-56.
233. Sandford A. J., Silverman E. K. Chronic obstructive pulmonary disease. 1: Susceptibility factors for COPD the genotype-environment interaction // Thorax. – 2002. – V. 57. - № 8. - P. 736-741.
234. Sandford A.J., Weir T.D., Pare P.D. et al. State of the Art. The genetics of asthma // Am. J. Respir. Crit. Care Med. – 1996. – V. 13. – P. 1749-1765.
235. Schwab M., Schaeffeler E., Klotz U. et al. CYP2C19 polymorphism is a major predictor of treatment failure in white patients by use of lansoprazole-based quadruple therapy for eradication of Helicobacter pylori // Clin Pharmacol Ther. – 2004. V. 76. – P.201-209.
236. Schwartz D.A., Freedman J.H. and Linney E.A. Environmental genomics; a key to understanding biology, pathophysiology and disease // Hum. Mol. Genet. – 2004. – V. 13. – P. 217-224.
237. Seidegard J., Vorachek W.R., Pero R.W. et al. Hereditary differences in the expression of human glutathione transferase activity on trans-stilbene oxide are due to a gene deletion // Proc. Nat. Acad. Sci. USA. – 1988. – V. 85. – P. 7293-7297.
238. Senak M., Pierzchalska M., Bazan-Socha S. et al. Enhanced expression of the leukotriene C(4) synthase due to overactive transcription of an allelic variant associated with aspirin-intolerant asthma // Am. J. Respir. Cell Mol. Biol. 2000. - № 23. – P. 290-296.
239. Sengler C. Lau S., Wan U. et al. Interactions between genes and environmental factors in asthma and atopy: new developments // Respir. Res. – 2002. – V. 3. № 7. – P. 7.
240. Shields P.G., Caporaso N.E., Falk R.T. et al. Lung cancer, race and CYP1A1 genetic polymorphism // Cancer Epidemiol. Biomarkers and Prev. – 1993. - № 2 – P. 481-485.
241. Shirakawa T., Enomoto T., Shimazu S. et al. The inverse assotiation between tuberculin responses and atopic disorder // Science. – 1997. – V. 275. – P. 77-79.
242. Shirakawa T., Li A., Dubowitz M. Association between atopy and variants of the beta subunit of the high-affinity immunoglobulin E receptor // Nat. Genet. – 1994. - 7. – P. 125-129.
243. Shirakawa T., Mao X.Q., Sasaki S. et al. Association between atopic asthma and a coding variant of Fc epsilon RI beta in a Japanese population [letter] // Hum. Molecular Genet. – 1996. - № 5. – P. 2068.
244. Sipes I.G., Gandolfi A.J. Biotransformation of toxicants // Casarett and Doull’s toxicology. – N. Y.: Macmillan Publishing Company. -1986. – P. 99-173.
245. Skadhauge L. R., Christens K., Kyvik K. O. et al. Genetic and environmental influence on asthma: a population-based study of 11,688 Danish twin pairs // Eur. Respir. J. – 1999. V. 13. – P. 8-14.
246. Sodhi C.P., Rana S.V., Mehta S. Study of oxidative stress in rifampicin induced hepatic injury in growing rats with and without protein energy malnutrition // Hum. Exp. Toxicol. 1997. - № 20. – P. 315-321.
247. Sodhi C.P., Rana S.V., Mehta S.K. et al. Study of oxidative stress in izoniazid induced hepatic injury in young rats with and without protein energy malnutrition // J. Biochem. Toxicol. – 1996. - № 11. – P. 139-146.
248. Spielman R.S., McGunis R.E., Ewens W.J. Transmission test for linkage disequilibrium: The insulin gene region and insulin-dependent diabetes-mellitus (IDDM) // Am. J. Hum. Genet. 1993. – V. 52. – P. 506-516.
249. Spitx M.R., Duphrone C.M., Detry M.A. et al. Dietary intake of isothiocyanates: evidence of a joint effect with glutathione S-transferase polymorphisms in lung cancer risk // Cancer Epidemiology, Biomarkers & Prevention. – 2000. - V. 9. – P. 1017-1020.
250. Spurdle A.B., Chen X., Abbazadegan M. et al. CYP17 promoter polymorphism and ovarian cancer risk // Int. J. Cancer. – 2000. - V. 86. – P. 436-439.
251. Stead W.W. Genetics and resistance to tuberculosis: could resistance be enhanced by genetics engineering? // Ann. Int. Med. – 1992. – V. 116. – P. 937-941.
252. Steele M.A., Burk R.F., Desprez R.M. et al. Toxic hepatitis with isoniazid and rifampicin – a meta analysis // Chest. – 1991. - № 99. – P. 465-471.
253. Sterk P.J., Fabbri L.M., Quanjer P.H. et al. Airway responsiveness // Eur. Respir. J. – 1993. – V. 6. Suppl. – P. 55-64.
254. Strachan D.P. Hayfever, hygiene, and household size // Biol.Med.J. – 1989. - V. 299. - P. 1259-1260.
255. Sugimura H., Suzuki I., Hamada G.S. et al. Cytochrome P450 IAI genotype in lung cancer patients and controls in Rio de Janeiro, Brazil // Cancer Epidemiol. Biomarkers and Prev. 1994. - № 3. – P. 145-148.
256. Summerhill E., Levitt S. A., Gidley H. et al. Beta(2)-adrenergic receptor Arg16/Arg16 genotype is associated with reduced lung function, but not with asthma, in the Hutterites // Am. J. Respir. Crit Care Med. – 2000. – V. 162. – P. 599-602.
257. Surry D.D., Neneses-Lorente G., Heavens R. et al. Rapid determination of rat hepatocyte mRNA induction potential using oligonucleotide probes for CYP1A1, 1A2, 3A, and 4A1 // Xenobiotica. - 2000. – V. 30. - № 5. – P. 441-456.
258. Tamer L., Calikoglu M., Ates N. A. et al. Glutathione S-transferase gene polymorphisms (GSTT1, GSTM1, GSTP1) as increased risk factors for asthma // Respirology. – 2004. – V. 9. - 4. – P. 493-498.
259. Tanaka K., Sugiura H., Uehara M. et al. Association between mast cell chymase genotype and atopic eczema: alone and those with atopic eczema and atopic respiratory disease // Clin. Exp. Allergy. – 1999. – V. 29. – P. 800-803.
260. The Collaborative Study in the Genetics of Asthma. A genome-wide search for allergic response (atopy) genes in three ethnic groups // Hum. Genet. – 2004. - V. 114. - P. 157-164.
261. The Collaborative Study on the Genetics of Asthma. A genome-wide search for asthma susceptibility loci in ethnically diverse populations. // Nat. Genet. – 1997. – V. 15. – P. 389-392.
262. Thomas N.S., Wilkinson J., Holgate S.T. The candidate region approach to the genetics of asthma and allergy. // Am. J. Respir. Crit Care Med. – 1997. – V. 156. – P. 144-151
263. To-Figueras J., Gene M., Gomez-Catalan J. Glutathione-S-transferase M1 and codon 72 p53 polymorphisms in a northwestern Mediterranean population and their relation to lang cancer susceptibility // Cancer Epidemiol. Biomark. Prev. – 1996. - V. 5. – P. 337-342.
264. Urs A. Molecular mechanisms of genetic polymorphisms of drug metabolism // Annu. Rev. Pharmacol. Toxicol. – 1997. – V. 37. – P. 169-196.
265. Van der Pouw Kraan T.C., Van Veen A., Boeije L.C. et al. An IL13 promoter polymorphism associated with increased risk of allergic asthma // Gene Immun. – 1999. - № 1. – P. 61-65.
266. Von Hertzen L., Klaukka T., Mattila H. et al. Mycobacterium tuberculosis infection and the subsequent development of asthma and allergic conditions // J. Allergy Clin. Immunol. 1999. – V. 104. - № 6. – P. 1211-1214.
267. Von Mutius E., Martinez F. D., Fritzsch C. et al. Prevalence of asthma and atopy in two areas of West and East Germany // Am. J. Respir. Crit. Care Med. – 1994. – V. 149. - № 2. – P. 358-364.
268. Watanabe J., Hayashi S., Kawajiri K. Different regulation and expression of the human CYP2E1 gene due to the RsaI polymorphism in the 5’-flanking region // J. Biochem. – 1994. – V. 116. – P. 321-326.
269. Watson M. A., Stewart R. K., Smith G. B. J. et al. Human glutathione S-transferase P1 polymorphism: relationship to lung tissue enzyme activity and population freguency distribution // Carcinogenesis. – 1998. – V. 19. – P. 275-280.
270. Waxman D.J. and Azaroff L. Phenobarbital induction of cytochrome P-450 gene expression. Biochem .J. – 1992. - V. 281. – P. 577-592.
271. Wen X., Wang J.S., Neuvonen J. et al. Isoniazid is a mechanism-based inhibitor of cytochrome P450 1A2, 2A6, 2C19 and 3A4 isoforms in human liver microsomes // Eur. J. Clin. Pharmacol. – 2002. – V. 57. № 11. – Р. 799 – 804.
272. Wheeler C., Guenthner T.M. Cytochrome P450-dependet metabolism of xenobiotic in human lung // J. Biochem. Toxicol. – 1991. – V. 6. – P. 163-169.
273. Whitlock J.P., Gelboin H.V. Aryl hydrocarbon (benzo[α]pyrene hydroxilase in rat liver cells in culture // J. Biol. Chem. - 1974. – V. 249. – P. 2616-2623.
274. Wilson M. H., Grant P.J., Hardine L.J. et al. Glutathione S-transferase M1 null genotype is associated with a decreased risk of myocardial infarction // The FASEB Journal. – 2000. - Vol. 14. – P. 791-796.
275. Wjst M., Fischer G., Immervoll T. et al. A genome-wide search for linkage to asthma. German Asthma Genetics Group // Genomics. – 1999. – V. 58. – P. 1-8.
276. Wright D.T., Cohn L.A., Li H. et al. Interactions of oxygen radicals with airway epithelium // Environ. Health Perspect. – 1994. – V. 102 (Suppl.10). – P. 85-90.
277. Xie H. G., Kim R. B., Stein C. M. et al. Genetic polymorphism of (S)-mephenytoin 4’-hydroxylation in populations of African descent // Br. J. Clin. Pharmacol. - 1999. – V. 48. - № 3. – P. 402-406.
278. Xu X., Kelsey K.T., Wiencke J.K. et al. Cytochrome P450 CYP1A1 MspI polymorphism and lung cancer susceptibility // Cancer Epidemiology, Biomarkers & Prevention. – 1996. - V. 5. – P. 687-692.
279. Yan L., Galinsky R. E., Bemstein J. A. et al. Histamine N-methyltransferase pharmacogenetics: assotiation of a common functional polymorphism with asthma // Pharmacogenetics. – 2000. - № 10. – P. 261-266.
280. Yang Y. S., Wong L. P., Lee T. C. et al. Genetic polymorphism of cytochrome P450 2C19 in healthy Malaysian subjects // Br. J. Clin. Pharmacol. – 2004. – V. 58. - № 3. – P. 332-335.
281. Yokouchi Y., Nukaga Y., Shibasaki M. et al. Significant evidence for linkage of mite-sensetive childhood asthma to chromosome 5q31-q33 near the interleukin 12 B locus by a genome-wide search in Japanese families // Genomics. – 2000. – V. 66. – P. 152-160.
282. Zhang YQ, Sun BY, Dai J.J. Studies on the genetic diathesis of asthma bronchial // Yi Chuan. – 2004. - 26. – V. 2. – P. 147-150.
283. Zhou W., Liu G., Thurston S.W. et al. Genetic polymorphisms in N-acetyltransferase-2 and microsomal epoxide hydrolase, cumulative cigarette smoking, and lung cancer // Cancer Epidemiology, Biomarkers & Prevention. – 2002. - V. 11. – P. 15-21.
БЛАГОДАРНОСТИ
Автор считает приятным долгом выразить огромную признательность своему руководителю – академику РАМН, профессору Пузыреву Валерию Павловичу за поддержку, чуткое отношение и неоценимую помощь в планировании, выполнении и обсуждении работы. Выражаю искреннюю благодарность сотрудникам лаборатории популяционной генетики ГУ НИИ МГ ТНЦ СО РАМН за предоставление материала и сопутствующей информации, а также за помощь в обсуждении полученных результатов.
Отдельную благодарность за предоставление клинического материала сотрудникам кафедры фтизиатрии Сибирского государственного медицинского университета (заведующий – член-корр. РАМН, профессор Стрелис А.К.) и сотрудникам кафедры факультетской педиатрии с курсом детских болезней лечебного факультета (заведующий – д.м.н., профессор Огородова Л.М.) Сибирского государственного медицинского университета, а также областного детского центра клинической иммунологии и аллергологии (Областная детская больница, г. Томск, главный врач – Сальников В.А.).
© 2010 Интернет База Рефератов