
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Контрольная работа: Отдел сине-зеленые водоросли
Контрольная работа: Отдел сине-зеленые водоросли
Отдел сине-зеленые водоросли – Cyanophyta
К данному отделу относят одноклеточные, колониальные и нитчатые водоросли, различно окрашенные в зависимости от соотношения пигментов, представленных кроме хлорофилла а и каротиноидов еще синими пигментами – фикоцианином и аллофикоцианином и красным фикоэритрином. Характерно также полное отсутствие жгутиковых стадий. Половой процесс у сине-зеленых водорослей не наблюдался.
Цитология. При рассматривании клетки сине-зеленых водорослей в оптическом микроскопе можно видеть, что окруженная оболочкой цитоплазма, как правило, лишена вакуолей с клеточным соком, окрашена в периферических частях и бесцветна в центре. Резкой границы между хроматоплазмой и центроплазмой нет. В центроплазме локализирована ДНК, что позволяет рассматривать ее как гомолог клеточного ядра, хотя ядерная оболочка и ядрышки отсутствуют. Кроме того, в цитоплазме находятся включения запасного характера и часто – полости, наполненные смесью газов, по составу близкой к воздуху. У одних видов псевдовакуоли встречаются на протяжении всей вегетативной жизни, у других – только на определенных стадиях развития.
В электронном микроскопе клетки сине-зеленых водорослей имеют следующее строение. Клеточная стенка состоит из четырех четко разграниченных слоев, обозначаемых Lx, L2, L3, L4. Непосредственно кнаружи от цитоплазматической мембраны расположен электронно-прозрачный слой за которым следует электронно-плотный слой, состоящий из муреина – основного компонента клеточной оболочки бактерий. У эукариотических. водорослей и грибов это вещество не обнаружено. Таким образом, и по химизму клеточной оболочки сине-зеленые водоросли, нередко называемые теперь сине-зелеными бактериями, более близки к бактериям, чем к эукариотическим водорослям. Именно этот муреиновый слой L2 определяет прочность стенки. Будучи изолирован, этот слой клеточной стенки способен сохранять форму всей клетки. Следующие за муреиновым слоем слои – электронно-прозрачный L3 и мембраноподобный L4 – образованы углеводами и в отличие от слоя L, гибкие, пластичные.
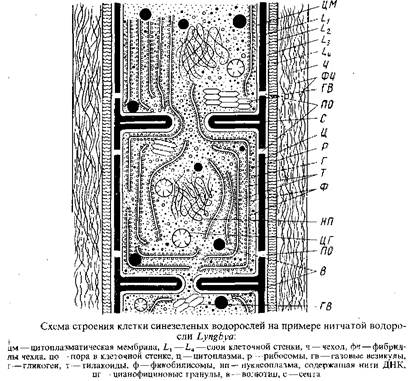
Обусловленная этими слоями пластичность клеточной стенки допускает изменения формы клетки. На уровне слоя L3 у осциллатории были обнаружены непрерывные параллельные ряды, по-видимому, протеиновых микрофибрилл, которые по спирали окружают трихом. Ориентация этих микрофибрилл совпадает с направлением вращения трихома во время его скольжения. Было высказано предположение, что волнообразные изгибы этих рядов микрофибрилл могут обусловливать скользящее движение трихома на твердом субстрате. Упомянутые четыре слоя наблюдаются в продольных стенках нитчатых сине-зеленых водорослей. Поперечные стенки, или септы, нитчатых сине-зеленых водорослей образованы только слоями L1 и L2. У одноклеточных форм наружные слои L3 и L4 образуются на септах только после начала разъединения клеток. Как в продольных стенках, так и в септах нитчатых сине-зеленых водорослей имеются поры, через которые соединяются цитоплазматические мембраны и протопласты соседних клеток; эти цитоплазматические тяжи называются микроплазмодесмами.
Подсчеты показали, что между двумя вегетативными клетками одного из видов анабенопсиса в септе может быть до 4000 микроплазмодесм.
У многих сине-зеленых водорослей клеточные стенки покрыты слизистым слоем, который может быть толстым и плотным и образует чехлы или капсулы, обычно заключающие несколько клеток, или же слизь представлена в виде тонкого жидкого слоя. Слизь предохраняет клетки от высыхания и, по-видимому, принимает участие в процессе скользящего движения. Тонкая структура слизи – фибриллярная.
Преимущественно в периферической цитоплазме – хроматоплазме – локализованы тилакоиды, способные иногда проникать во все части клетки. Тилакоиды не отграничены от цитоплазмы мембранами, как у других хлорофиллоносных растений, т.е. истинные хлоропласты, одетые оболочкой, здесь отсутствуют. Периферически расположенные тилакоиды обычно ориентированы параллельно продольной клеточной стенке, но иногда они расположены перпендикулярно к продольной стенке. Изредка тилакоиды рассеяны по всей клетке.
Тилакоиды сине-зеленых водорослей никогда не образуют групп, как это свойственно эукариотическим водорослям, за исключением красных, где тилакоиды также расположены одиночно. Добавочные пигменты сине-зеленых водорослей в форме гранул – фикобилисом локализованы на поверхности тилакоидов.
Центр клетки, обычно свободный от тилакоидов, занят нуклеоплазмой, не отграниченной от остальной цитоплазмы ядерной оболочкой» В нуклеоплазме находятся фибриллы ДНК. Для ядерного материала сине-зеленых водорослей, как и для бактерий и фагов, характерно отсутствие гистонов, чем они отличаются от остальных эукариотических организмов, ядерная субстанция которых содержит помимо ДНК, ядерный белок.
В цитоплазме, не занятой тилакоидами и нуклеоплазмой, находятся рибосомы и запасные вещества: гликоген, волютин, цианофициновые гранулы.
Газовые вакуоли, рассеянные по всей клетке или располагающиеся у поперечных перегородок, состоят из тесно упакованных, наподобие сот, полых, одетых мембраной субъединиц газовых везикул. Эти везикулы имеют форму полых цилиндрических трубок с коническими шапочками у концов. Они могут быть изолированы из клетки без изменения их формы. Подвергнутые определенному давлению цилиндрические части газовых везикул спадаются, при этом конические шапочки могут отламываться. Химические анализы изолированных газовых везикул показали, что их мембраны отличаются от типичной элементарной мембраны отсутствием липидов. Мембраны газовых везикул состоят только из белков.
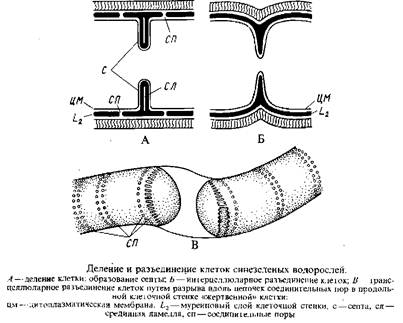
Размножение. Перед клеточным делением количество ДНК удваивается, а по мере деления клетки удвоенное количество ДНК разделяется пополам. Клетки сине-зеленых водорослей делятся таким образом, что на боковой стенке их возникает кольцевая складка, образованная цитоплазматической мембраной и внутренними слоями клеточной оболочки. Разрастаясь в центростремительном направлении, yia складка смыкается наподобие ирисовой диафрагмы микроскопа, формируя поперечную перегородку – септу, перерезающую тилакоиды и содержимое клетки.
В середине поперечно перерезанной септы в процессе утолщения муреинового слоя сравнительно рано становится различимым слой более низкой плотности – срединная ламелла. У одноклеточных форм и многих ностокальных, у которых образование клеточной перегородки сопровождается появлением более или менее глубокой перетяжки между дочерними клетками, клетки разъединяются путем центрипетального врастания в плоскость срединной ламеллы между слоями муреина (L2) наружных слоев клеточной оболочки. Таким образом, у многих ностокальных разрыв трихома при образовании гормогониев происходит между клетками – интерцеллюларно. Совсем иначе разъединяются клетки у многих осциллаториальных и сцитонемовых. Хотя муреиновые слои зрелой септы разъединены срединной ламеллой, расщепления септы и расхождения клеток в этой области не происходит, так как муреиновые слои закреплены у периферического утолщения края диска септы. Слабым местом в муреиновом одеянии клеток у форм с нерасщепляющейся септой, по-видимому, является набор многочисленных мелких соединительных пор, расположенных по окружности по обе стороны от септы – вдоль соединения ее с продольными стенками. Разрыв трихомов при образовании гормогониев происходит вдоль соединительных пор – поперек клеток – трансцеллюларно и связан с гибелью клетки, «приносимой в жертву» – «жертвенной клетки». При этом ненарушенные септы между «жертвенной клеткой» и двумя соседними с ней клетками становятся стенками конечных клеток сформировавшихся гормогониев. Остатки клеточных стенок «жертвенных клеток» могут служить доказательством того, что разрыв произошел не благодаря расщеплению септ между живыми клетками, а трансцеллюларно. Такие остатки оболочек отмерших клеток, по которым происходит разрыв, часто можно видеть в оптическом микроскопе на концах трихомов осциллаторий, Как интерцеллюларным, так и трансцеллюларным разрывам способствуют многократные изгибы трихомов.
У одних нитчатых сине-зеленых водорослей все клетки нити одинаковы – это гомоцитные талломы у других нити состоят из вегетативных клетки гетероцист, нередко наблюдаются и споры – гещероцитные формы. Гетероцисты и споры – это специализированные клетки, образующиеся путём «дифференцировки вегетативных клеток. Один из наиболее заметных признаков гетероцист – их сильно утолщенные стенки. Клеточные стенки гетероцист, как и вегетативных клеток, состоят из слоев Lx–L4, но кнаружи от них развиваются еще три специальных слоя оболочки гетероцист.
Непосредственно кнаружи от клеточной стенки располагается пластинчатый слой, особенно толстый у глубокой перетяжки между гетероцистой и соседней вегетативной клеткой, где имеется поровый канал – «пора», пересеченная септой, пронизанной микроплазмодесмами, посредством которых сообщаются гетероциста и вегетативная клетка. Кнаружи от пластинчатого слоя находится гомогенный слой, который без резкой границы переходит в самый наружный фибриллярный слой. Благодаря толстой обвертке, образующейся кнаружи от клеточной оболочки, гетероцисты выдерживают механические воздействия, разрушающие более тонкостенные вегетативные клетки.
Другой
признак зрелых гетероцист, легко наблюдаемый в оптический микроскоп, – гомогенность
клеточного содержимого, которая обусловлена исчезновением гранулярных
включений, характерных для вегетативных клеток. Ни волютина, ни цианофициновых
зерен в полностью развитых гетероцистах нет. Единственные гранулярные
структуры, которые можно различить на электронных микрофотографиях зрелых
гетероцист, – рибосомы. Газовые вакуоли, если они были в вегетативных клетках,
при дифференцировке их в гетероцисты также исчезают.
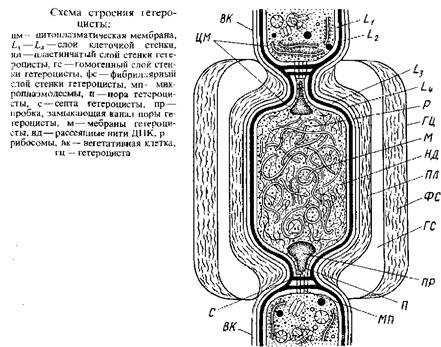
Наконец, дифференцировка гетероцист сопровождается реорганизацией мембранной системы клетки: тилакоиды разрушаются и формируются новые плотно упакованные мембраны. Преобразование мембранной системы сопряжено с изменениями в пигментном составе: в гетероцистах обнаруживаются хлорофилл и каротиноиды, но почти нет фикоцианина, аллофикоцианина и фикоэритрина.
Фибриллы ДНК, в вегетативных клетках обычно сконцентрированные в нуклеоплазме, в гетероцистах рассеяны по всей цитоплазме.
В многочисленных исследованиях было показано, что образование гетероцист, как и процесс фиксации атмосферного азота, ингибируется присутствием в питательной среде связанного азота, особенно аммонийного. Исходя из этих данных стало возможным при выращивании на средах, содержащих связанный, особенно аммонийный азот, искусственно получать трихомы без гетероцист у родов, в норме характеризующихся наличием гетероцист, например у анабен, ностоков и др. При перенесении такого безгетероцистного материала в среду, свободную от связанного азота, можно проследить все стадии развития гетероцист из вегетативных клеток. Дифференцировка гетероцист начинается с образования самого наружного фиброзного слоя ее оболочки кнаружи от клеточной стенки дифференцирующейся вегатативной клетки. Образование фиброзного слоя и следующего гомогенного слоя сопровождается разрушением гранулярных цитоплазматических включений и тилакоидов. На конечной стадии развития гетероцист откладывается самый внутренний пластинчатый слой оболочки гетероцисты, особенно толстый вокруг порового канала, и формируется развитая мембранная система, по-видимому, путем соединения мелких пузырьков, возникающих на местах предшествующих тилакоидов.
Давно известно, что у таких родов, как анабена, носток, характеризующихся интеркалярными гетероцистами, в гетероцисты превращаются не любые вегетативные клетки, а лишь некоторые, расположенные на определенном расстоянии друг от друга. Сравнительно недавно было высказано предположение, что гетероцисты анабены выделяют ингибитор, подавляющий дифференцировку вегетативных клеток в гетероцисты. Последние возникают из вегетативных клеток, расположенных вне ингибирующих зон, окружающих ранее образовавшиеся гетероцисты.
В ряде работ эту гипотезу удалось подтвердить экспериментально. Так, Д. Тивари из дикого штамма Nostoc linckia, способного осуществлять процесс азотфиксации, выделил мутанты, не фиксирующие атмосферный азот, по сохранившие возможность осуществлять развитие, ведущее к дифференцировке гетероцист. На средах, не содержащих азота, у этих штаммов появлялись иптеркалярные гетероцисты. Однако в отличие от дикого исходного штамма вегетативные клетки по соседству с возникшими гетероцистами также превращались в гетероцисты, что приводило к образованию цепочек гетероцист. Напрашивается вывод, что ингибитор – это дериват какого-то непосредственного продукта азотфиксации. Он максимально концентрируется вокруг функционирующих гетероцист, регулируя распределение гетероцист вдоль нити. У других родов гетероцисты занимают терминальное положение в трихоме; бывают и латеральные гетероцисты.
У очень многих гетероцитных форм, в зрелом состоянии характеризующихся интеркалярными гетероцистами, их молодые проростки имеют терминальные гетероцисты.
Относительно функции гетероцист высказывались разные предположения. У гетероцитных форм распад нитей на отдельные участки, из которых развиваются новые талломы, обычно происходит по гетероцистам, соответственно им приписывалась и известная роль в вегетативном размножении. Наблюдаемые иногда случаи деления содержимого гетероцист позволяли предположить их первоначальную роль в качестве клеток, предназначенных для размножения, И действительно, при культивировании в определенных условиях описано довольно много фактов прорастания гетероцист. Так, у Anabaena cylindrica, выращенной в безазотистой среде, при перенесении ее на среду с хлоридом аммония 3–10% гетероцист прорастали. У неспорулирующего мутанта одного из видов Gloeotrichia при культивировании в среде с хлоридом аммония прорастало 83% гетероцист. Сравнительно недавно было даже высказано предположение, что исчезновение гетероцист, наблюдаемое на средах, в состав которых входит связанный азот, объясняется как раз их прорастанием. Однако в естественных условиях прорастание гетероцист – редкое явление. Против их роли в качестве клеток бесполого размножения свидетельствует не только то, что они лишены запасных продуктов, которые могли бы поддержать начальные стадии прорастания, но и то, что при отделении их от нити, они, как правило, быстро разрушаются, возможно, оттого, что в отличие от спор не защищены со всех сторон оболочкой. Согласно последним данным в гетероцистах происходит фиксация атмосферного азота в аэробных условиях.
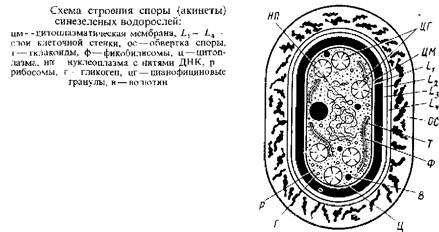
Другими специализированными клетками, которые также образуются путем дифференцировки вегетативных клеток, являются споры – акинеты. Это обычно более крупные, чем вегетативные, клетки, толстостенные, причем споровый покров окружает акинету полностью в отличие от гетероцист, которые через поровый канал сохраняют контакт с соседними вегетативными клетками. При дифференцировке спор из вегетативных клеток заметно утолщается L2 слой и, кроме того, кнаружи от клеточной оболочки образуется широкая обвертка. В отличие от гетероцист зрелые споры переполнены запасными гранулярными включениями, особенно цианофициновыми зернами. Расположение и структура тилакоидов в спорах те же, что и в вегетативных клетках. Содержание ДНК в спорах заметно возрастает, иногда в 20–30 раз по сравнению с содержанием ее в вегетативных клетках. Как и в случае гетероцист, вегетативные клетки, превращающиеся в споры, утрачивают газовые вакуоли.
Спорообразование у сине-зеленых водорослей стимулируется разными факторами среды, но особенно фосфором. Споры могут выдерживать высыхание и другие неблагоприятные условия. Они могут прорастать сразу же после образования, без какого-либо периода покоя, при помещении в благоприятные условия. При отсутствии таких условий споры долго сохраняют жизнеспособность. Например, споры Anabaena spiroides, A. flos-aquae, Aphanizomenon flos-aquae, выделенные из донных осадков, возраст которых составлял от 18 до 64 лет, хорошо прорастали на минеральной среде в лабораторных условиях. При прорастании споры образуется один проросток, освобождающийся через разрыв оболочки.
Большинство одноклеточных и колониальных сине-зеленых водорослей размножаются посредством деления клеток пополам, у некоторых – посредством мелких клеток – «гонидий», образующихся или внутри материнской клетки, или отшнуровывающихся от верхушки материнской клетки. Подавляющее большинство нитчатых сине-зеленых водорослей размножается посредством гормогоииев – участков, па которые распадаются нити.
Синезеленые водороли подразделяют на три класса: хроококкофициевые, хамесифонофициевые и гормогониофициевые.
Класс xроококкофициевые – chroococcophyceae
Объединяет колониальные, реже одноклеточные формы, причем клетки, как правило, не дифференцированы на основание и вершину. Размножение – делением клеток пополам. Если клетки после деления не расходятся, то обычно возникают слизистые колонии, форма которых определяется как формой самих клеток, так и способом их деления, в одних случаях приводящим к возникновению объемных колоний, в других – плоских, пластинчатых.
Род микроцистис, широко распространенный в пресноводном планктоне, где, развиваясь в массе, вызывает «цветение воды», характеризуется объемными сферическими или неправильной формы слизистыми колониями, образованными шаровидными клетками. Некоторые виды токсичны.
Род мерисмопедия, обычный в пресных водах в планктоне и среди других водорослей, представлен плоскими, в виде табличек, колониями, образованными шаровидными клетками.
Класс хамесифонофициевые – chamaes1phonophyceae
Сюда относятся эпифитные одноклеточные водоросли, дифференцированные на вершину и основание, которым они прикрепляются к субстрату, а также нитчатые формы, сложенные из изолированных толстостенных клеток. Размножение – посредством эндоспор и экзоспор.
Класс гормогониофициевые – hormogoniophyceae
Этот класс объединяет нитчатые многоклеточные синезеленые водоросли, у которых протопласты соседних клеток сообщаются посредством плазмодесм. Такие нити носят специальное название трихом. Размножение посредством гормогониев, которые обычно определяются как участки трихомов, способные к активному движению и прорастанию в новые особи. У многих представителей известны споры. Они прорастают также гормогониями, которые в отличие от вторичных гормогониев, развивающихся из взрослых трихомов, называются первичными. Из входящих сюда порядков рассмотрим осциллаториальные, ностокальные и стигонемальные.
Порядок осциллаториальные – oscillatoriales
Порядок объединяет нитчатые гомоцитные формы. Представителем может служить род осциллатория. Его виды часто образуют сине-зеленые пленки, покрывающие влажную землю, подводные предметы, или плавают в виде толстых кожистых лепешек на поверхности стоячих водоемов. Длинные нити осциллатории сложены из цилиндрических клеток, совершенно одинаковых, за исключением верхушечных, которые по форме могут несколько отличаться от остальных. Рост происходит в результате поперечных делений клеток. Для нитей характерно своеобразное колебательное движение, сопровождающееся вращением нити вокруг собственной оси и ее поступательным движением. Размножение – путем траисцеллюларного распада трихомов по «жертвенным» клеткам на более короткие подвижные участки, часто определяемые как гормогонии. Они вырастают в новые нити.
Во многих случаях трансцеллюларный распад происходит по уже мертвым клеткам – некридиям.
Близок к осциллатории род спирулниа, нити которой скручены в правильную спираль. Некоторые виды спирулины, например S. maxima, содержат много протеинов и с давних времен используются в пищу населением некоторых районов Африки. В последние годы в ряде стран, в том числе в СССР, стали заниматься массовой культурой видов спирулины.
Порядок ностокальные – Nostocales
Порядок объединяет водоросли с гетероцитными неразветвленными нитями или нитями с ложным ветвлением.
Род анабена обычно одиночные или собранные в неправильные скопления нити. Нити симметричные, одинаковой ширины на всем протяжении, состоят из округлых или бочонкообразных вегетативных клеток и промежуточных гетероцист. Нити прямые или изогнутые. Виды анабены встречаются как в бентосе, так и в планктоне. У планктонных, видов клетки содержат газовые вакуоли. Размножение – посредством подвижных нитей, часто называемых гормогониями, образование которых сопровождается интерцеллюларными разрывами обычно между вегетативными клетками. Таким образом, на протяжении активно ползающих «гормогониев» анабен имеются интеркалярные гетероцисты родительского трихома, чем они отличаются от гормогониев других родов, как правило, гомоцитных. Реже у тех же видов происходит интерцеллюларный разрыв между вегетативными клетками и гетероцистами, сначала еще живыми и лишь позднее отмирающими, возможно, в результате нарушения микроплазмодесм, связывающих гетероцисты с соседними вегетативными клетками. «Гормогонии» растут только за счет поперечных делений клеток. Большинство видов имеет споры, обычно резко отличающиеся по форме и размерам от вегетативных клеток. Их прорастание сопровождается только поперечными делениями клеток. Первые формирующиеся в проростке гетероцисты часто термальные. Для ряда видов доказана способность к фиксации атмосферного азота.
Род носток характеризуется слизистыми или студенистыми колониями различных размеров и форм: от микроскопически мелких до крупных, достигающих величины сливы; от сферических или эллипсоидальных до неправильно распростертых и нитевидных. Слизь разной консистенции содержит массу извитых нитей или беспорядочно перепутанных, или расходящихся более или менее радиально из центра колонии; трихомы похожи на трихомы анабены; наряду с вегетативными клетками содержат интеркалярные гетероцисты. Размножение – посредством гормогониев. Последние возникают в результате интерцеллюлярных разрывов между вегетативными клетками и гетероцистами; таким образом, гормогонии всегда гомоцитные. Отмирание гетероцист, по-видимому, предшествует разрыву; выпадают из трихома лишь пустые оболочки гетероцист, в результате чего трихомы распадаются на гормогонии. Это наблюдается у таких видов, как Nostoc punctiforme, N. pruniforme и др. Образование гормогониев сопровождается внутриклеточными перестройками, внешне выражающимися в том, что в большинстве случаев изменяются размеры и форма клеток. Очень часто у бентосных форм, в вегетативном состоянии лишенных газовых вакуолей, эти вакуоли появляются в клетках гормогониев. Гормогонии приобретают подвижность и покидают материнскую колонию, слизь которой к этому времени расплывается, а если колония покрыта плотным наружным слоем – перидермом, то этот слой разрывается. Обычно гормогонии служат и для распространения вида благодаря собственному скользящему движению и вследствие лучшей плавучести, обусловленной развитием в клетках газовых вакуолей. После некоторого периода движения прямые гормогонии останавливаются, теряют газовые вакуоли и прорастают в спирально извитые нити. При этом клетки гормогония делятся косыми или продольными перегородками. Во втором случае сначала возникает двурядная нить, а затем при разъединении определенных клеток этой нити формируется уже зигзагообразная нить, свойственная ностокам. Обычно конечные клетки молодой нити превращаются в гетероцисты, выделяется обильная слизь и таким образом возникает молодая колония, в которой нити дальше растут только за счет поперечных делений клеток. У N. punctiforme - вида, лишенного крепкого перидерма, дочерние колонии возникают за счет развития многих гормогониев, У сферических ностоков с крепким перидермом дочерние колонии образуются в результате прорастания только одного гормогония. Длинные гормогонии ностоков могут непосредственно прорастать в извитые трихомы, но в некоторых случаях гормогонии претерпевают предварительную фрагментацию на короткие отрезки; фрагментация сопровождается многократными изгибами длинных гормогониев. Эти изгибы способствуют интерцеллюларным разрывам. Фрагментация заметно повышает энергию размножения. Это относится и к самому крупному сферическому ностоку – N. pruniforme, развивающемуся в наших северных водоемах в колоссальных количествах. В ряде случаев у сферических ностоков может быть подавлена подвижность гормогониев, которые не покидают слизи материнской колонии и развиваются внутри нее с образованием эндогенных колоний, позднее освобождающихся в результате разрушения перидерма материнской колонии. Иногда у ностоков наблюдаются ветвящиеся гормогонии, которые прорастают, как и обычные неразветвленные гормогонии. Сферические ностоки нередко размножаются почкованием колоний. Помимо гормогониев, у многих ностоков образуются споры, обычно мало отличающиеся по форме и размерам от вегетативных клеток. Нередко все вегетативные клетки превращаются в споры, остающиеся связанными в цепочки. При прорастании их оболочки разрушаются, и проросток, еще не поделившийся или уже многоклеточный, освобождается. Как у вторичных, так и у первичных гормогониев иногда наблюдается полная потеря подвижности, и они прорастают на месте образования внутри оболочек спор. У N. linckia концы проростков, развившихся при прорастании таких неподвижных первичных гормогониев, соединяются с образованием как бы омоложенного нового трихома, и на месте спороносной колонии, содержащей цепочки спор, формируется вегетативная. Самый крупный N. pruniforme, а также носток с колониями в виде слизистых тяжей, в которых нити расположены более или менее параллельными рядами, съедобны. Ностоки способны фиксировать атмосферный азот.
У ряда других ностокальных трихомы асимметричны. Например, у рода калотрикс трихомы бичевидные: на расширенном конце дифференцируется базальная гетероциста, противоположный конец утончается и часто заканчивается многоклеточным волоском из отмерших клеток. Трихом одет чехлом и может ветвиться. Ветвление ложное и обусловлено тем, что на протяжении трихома в результате отмирания промежуточной клетки происходит разрыв. Расположенный ниже разрыва участок, прорывая чехол, продолжает расти вбок, образуя ветвь. Конечная клетка верхнего фрагмента может превратиться в базальную гетероцисту или же базальный конец верхнего фрагмента может дать вторую боковую ложную ветвь. В первом случае ложное ветвление одиночное, во втором – двойное. При размножении С. stellaris и других видов волоски сбрасываются, клетки трихома, начиная с его верхушки, переполняются газовыми вакуолями, несколько меняются их форма и размеры. Эти изменения не затрагивают лишь несколько базальных клеток. Возникает длинная гомоцитная нить – прогормогоний, который обычно покидает чехол материнского трихома. Прогормогоний способен к движению, но не к непосредственному прорастанию в новые трихомы. Прогормогоний выполняют функцию распространения вида, которая осуществляется не столько благодаря собственному активному скользящему движению, сколько за счет газовых вакуолей, переполняющих их клетки и способствующих их лучшей плавучести. Такие прогормогонии закономерно претерпевают фрагментацию на более короткие участки гормогонии. также сначала подвижные и способные прорастать с образованием трихома. Фрагментация активная, сопровождается многократными изгибами прогормогония и может продолжаться несколько часов. Иногда подвижность прогормогония подавлена и распад его на фрагменты происходит пассивно внутри влагалища материнского трихома, из которого выходят уже подвижные фрагменты – гормогонии. При прорастании гомоцитных гормогониев возникают асимметричные, гетероцитные, с терминальной гетероцистой на расширенном конце трихомы. У некоторых видов калотрикса известны споры.
У родов ривулярия и глеотрихия талломы студенистые: в слизи, расходясь из центра по радиусам, располагаются асимметричные, как у калотрикса, нити. К центру колонии обращены расширенные концы с базальной гетероцистой, к периферии – волоски. Размножение ривулярии и глеогрихии – посредством гормогониев. У всех видов глеотрихии известны споры, у ривулярии споры отсутствуют. Размножение и развитие колоний подробно изучено у многих видов глеотрихии. В вегетативных колониях из трихомов, как и у калотрикса, сначала формируются длинные симметричные гомоцитные черные прогормогонии. Они покидают слизь материнской колонии, образуют плотные пучки и всплывают на поверхность воды. В таких пучках прогормогопиев вскоре наступает активная, сопровождающаяся коленообразными изгибами ломка их на более короткие фрагменты. Фрагментация, по-видимому, трансцеллюлярная, обычно происходит одновременно во всех прогормогониях, слагающих снопик, и минут через 10–20 на его месте свободно лежат короткие фрагменты и вытолкнутые «жертвенные» клетки. Через некоторое время от и клетки теряют содержимое и опускаются на дно сосуда. Распад прогормогониев продолжается до образования гормогониев – коротких участков, при дальнейшем развитии которых возникает только один трихом, или образуются более длинные вигормогонии, дающие два трихома. В первом случае, характерном для Gloeotrichia pisuni, совершенно прямые и гомоцитные вначале гормогонии, сложенные из одинаковых черных клеток, останавливаются и начинают медленно изгибаться дугой, В них происходит дифференцировка клеток: па одном конце намечается будущая гетероциста, на противоположном конце – едва заметное утончение и удлинение клеток. По мере дальнейшего развития асимметричного трихома газовые вакуоли из клеток исчезают. Во втором случае, свойственном Gl. echiiiulata, бигормогонии сначала образуют симметричный, утончающийся к обоим концам трихом, впоследствии распадающийся на два асимметричных трихома благодаря формированию двух средних гетероцист и последующему разрыву между ними. Поскольку этот вид планктонный, газовые вакуоли в клетках сохраняются. По мере дифференцировки клеток трихомов выделяется слизь, в которой трихомы располагаются более или менее по радиусам. Таким образом формируются молодые колонии, как правило, за счет развития многих гормогониев, чему способствует то, что с самого начала прогормогоний собраны в плотные пучки. У глеотрихии длинная, с зернистым содержимым спора образуется за счет слияния нескольких клеток, расположенных над базальной гетероцистой. К концу вегетационного периода все нити в колонии образуют споры, вегетативные клетки отмирают и в слизи остаются только споры; у шаровидных видов споры расходятся из центра колонии по радиусам. При помещении таких спороносных колоний в благоприятные условия споры более или менее одновременно прорастают. Содержимое их делится с образованием длинных симметричных многоклеточных нитей прогормогониев, в клетках которых обычно появляются обильные газовые вакуоли. Оболочка на верхушке споры разрывается и более легкие, чем вода, обычно соединенные пучками прогормогоний всплывают на ее поверхность, где и претерпевают дальнейшее развитие, совершенно такое же, как это было описано для прогормогониев, образующихся из взрослых вегетативных колоний.
Порядок стигонемальные – Stigonematales
Этот порядок характеризуется гетероцитными нитями с настоящим ветвлением. При настоящем – истинном – ветвлении отдельные клетки трихома образуют выступы, которые отделяются перегородкой и затем, повторно делясь поперечными перегородками, развиваются в ветви. Примерами могут служить роды стигонема и мастигокладус. Нити стигонемы обладают верхушечным ростом. Несколько отступая от верхушки ветвей, клетки претерпевают продольные деления, и нить становится многорядной. Мастигокладус с единственным видом М. laminosus имеет сложный ветвящийся гетероцитный таллом и является типичной термальной водорослью, распространенной по всему свету. Образование гормогониев, в которые превращались концы боковых ветвей, у этой термофильной водоросли наблюдается при 40–45°С, хотя талломы не теряют жизнеспособности и даже обнаруживают слабый рост при температурах ниже 30°С. В стареющих культурах возникают споры, расположенные цепочками. У них нет периода покоя и в благоприятных условиях они часто прорастают in situ, нередко имитируя при этом боковые ветви.
Таким образом, процесс образования гормогониев у разных представителей существенно различается. Лишь у осциллаториальных гормогонии отвечают вышеприведенному определению их как участков трихомов, способных к активному движению и прорастанию в новые особи. Разрыв гомоцитных трихомов осциллатории на гормогонии, отличающиеся от зрелых особей только меньшей длиной, можно рассматривать как типичный пример вегетативного размножения – распада таллома на отдельные участки без каких-либо перестроек в содержимом клеток. У представителей вышестоящих порядков гормогониофициевых – ностокальных и стигонемальных, имеющих гетероцитные трихомы, образование гормогониев нельзя свести к простому разрыву трихомов на отдельные участки. Здесь формирование гормогониев – гораздо более сложный процесс, как правило, сопровождающийся внутриклеточными перестройками, о которых пока мало известно. Обычно внешне это выражается в появлении в клетках гормогониев новых органелл, отсутствующих в материнском трихоме, – газовых вакуолей, а также в значительном изменении формы и размеров клеток. Здесь гормогонии, почти всегда гомоцитные и симметричные, резко отличаются от гетероцитных, часто асимметричных зрелых трихомов. Оба признака гормогониев осцилляториальных–подвижность и способность к прорастанию в новые трихомы – часто бывают разобщены в прогормогониях, возникающих у многих представителей ностокальных при их размножении и часто называемых гормогониями. В связи с этим необходимо более резко разграничивать образование гормогониев от случайных разрывов материнских трихомов на более короткие участки и согласиться с несколько иным, чем было приведено выше, определением самого понятия «гормогонии». Так, Р. Ринпка и др. определяют гормогонии как трихомы, отличающиеся от родительских отсутствием гетероцист, а также одним или совокупностью таких признаков, как быстрое скользящее движение, иные размеры и форма клеток, присутствие газовых вакуолей в клетках. Этому определению отвечают структуры, служащие для размножения родов носток, калотрикс, глеотрихия, мастигокладус и др. Размножение же осциллаториальных и анабен следует рассматривать как случайный разрыв родительского трихома на участки, отличающиеся от трихома только меньшей длиной. Основное различие между родами носток и анабена указанные авторы видят именно в способах размножения: в наличии гормогониев у ностока и отсутствии гормогониев у анабен. Следует, однако, добавить, что фрагменты, на которые распадаются трихомы анабен, отличаются от материнских подвижностью и часто присутствием газовых вакуолей. Что касается критериев для разграничения оодов анабена и носток, то полезен и такой признак, как характер деления клеток проростков, развивающихся из спор: у анабены происходят только поперечные деления клеток, у разных видов ностока – косые и продольные.
Подвижность прогормогониев и гормогониев сильно варьирует у разных видов и штаммов и зависит от условий внешней среды. Так, потребность в свете для движения гормогониев у разных гормогониофициевых неодинакова. Здесь можно выделить по крайней мере три группы видов:
1. Виды и штаммы, гормогонии и прогормогоний которых одинаково подвижны как на свету, так и в темноте; некоторые штаммы jV. punciifortne, Calothrix elenkini, С. braunii.
2. Виды, гормогонии которых для приобретения подвижности нуждаются в свете. Однако при добавлении в среду глюкозы движение гормогониев в темноте не отличалось от движения их на свету.
3. Виды, гормогонии которых обнаруживают подвижность только на свету, в темноте движения не наблюдается даже при добавлении в среду углеводов. Сюда в первую очередь относится термальная водоросль Mastigocladus laminosus, для образования гормогониев которой необходимы высокие температуры. Кроме того, для приобретения подвижности и миграции по поверхности агара образовавшихся гормогониев необходим свет. В темноте при оптимальных температурах гормогонии остаются неподвижными. Добавление же в среду различных доз глюкозы не оказывало никакого воздействия на подвижность гормогониев. Движение начиналось только после выставления культур на свет: через непродолжительный промежуток времени наблюдалась миграция гормогониев от места посева по поверхности агара. На свету глюкоза оказывает некоторый стимулирующий эффект на интенсивность движения гормогониев, которые расползаются от посева на большую дистанцию, чем в отсутствие глюкозы.
Свет необходим также для приобретения подвижности теми участками, на которые распадаются трихомы анабен.
В ряде случаев подвижность гормогониев может быть утрачена полностью.
При прорастании гормогониев во взрослые трихомы ход их развития в значительной мере обратный ходу развития гормогониев из трихомов, г. е. из гомоцитной нити при подходящих внешних условиях в конце концов развивается нить, свойственная данному роду.
В процессе индивидуального развития гормогониофициевых водорослей они проходят ряд неодинаковых морфологических стадий или статусов, сходных со зрелыми особями других родов. Так, все гормогониофициевые водоросли в состоянии гормогониев отвечают диагнозу рода осциллатория.
При дальнейшем развитии разные роды различаются, по статусам, и число их тем больше, чем сложнее организация таллома данного рода. Так, у мастигокладуса со сложным гетероцитным талломом с дифференцировкой вегетативных клеток и разными типами ветвления в жизненном цикле удается установить десять статусов.
На разных ростовых фазах или статусах гормогониофициевые водоросли могут задерживать свое развитие, вегетировать длительное время и даже размножаться посредством гормогониев.
Это послужило причиной того, что отдельные статусы в цикле развития ностокальных водорослей неоднократно описывались как самостоятельные таксоны. Поэтому система гормоониофициевых водорослей нуждается в пересмотре на основе уточнения их онтогенеза. Онтогенез удобнее всего изучать в условиях культуры. Однако при этом не следует игнорировать наблюдения в естественных местообитаниях, как это делают некоторые бактериологи. Так, Р. Риппка и др., изучая в культуре сине-зеленые водоросли глеотрихию и ривулярию, постоянно существующие в природе в виде слизистых колоний, не образующие их при некоторых условиях культивирования, приходят к выводу об упразднении этих родов. Для цианобактерий с асимметричными трихомами и базалькой гетероцистой они оставляют один род калотрике.
Сине-зеленые водоросли – очень древние, известны из докембрийских отложений. Распространены они повсеместно, растут там, где не могут обитать никакие другие растения и первыми заселяют вновь образующиеся поверхности Земли, например вулканические острова. Большинство сине-зеленых водорослей встречается в пресных водах, меньше – в море. В планктоне прудов, озер, медленно текущих рек виды родов микроцистис, анабена, глеотрихия часто вызывают «цветение» воды.
Сине-зеленые водоросли имеют большое значение в жизни человека, как положительное, так и отрицательное.
Отдел прохлорофитовые водоросли – prochlorophyta
Это новый отдел растительного царства, предложенный Р. Левиным для одноклеточных сферических водорослей, размножающихся путем деления клетки посредством экваториальной перетяжки, относимых к новому роду прохлорон – Prochloron. Эти водоросли живут на поверхности или в клоаке некоторых асцидий. Замечательная особенность их – сочетание прокариотной организации клетки с пигментным составом, свойственным зеленым водорослям: доказано присутствие у прохлорона хлорофиллов а и отсутствие фикобилиновых протеиновых пигментов, характерных для сине-зеленых водорослей. Фотосинтетические ламеллы состоят большей частью из двух сближенных тилакоидов с гладкими мембранами, лишенными фикобилисом.
© 2010 Интернет База Рефератов