
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Контрольная работа: Сравнительный обзор кожных покровов у хордовых
Контрольная работа: Сравнительный обзор кожных покровов у хордовых
Контрольная работа
по зоологии позвоночных
Тема:
Сравнительный обзор кожных покровов у хордовых
План работы
Вступление
1.Сравнительный обзор кожных покровов у хордовых животных
2. Особенности строения кожных покровов и их производных с учетом различных условий жизни хордовых животных
2.1 Особенности строения и образования чешуи;
2.2 Особенности строения и образования пера,
2.3 Особенности строения шерсти (волосы) и другие образования кожи.
2.4 Кожные железы наземных позвоночных. Функции, особенности строения
Заключение
Вступление
Необходимое условие эволюционного развития − усложнение организации в одном направлении и упрощение в другом, т. е. сочетание в едином процессе прогрессивных и регрессивных явлений. Позвоночные объединены общностью морфофизиологической организации. Во всех системах органов этих животных можно проследить черты преемственных изменений в связи с эволюционным преобразованием органов. Кожные покровы представляют собой весьма важную в функциональном отношении систему. Кожа и мускулы оформляют тело животного с поверхности, придают ему форму и удерживают все внутренние органы. Кожные покровы защищают тело от внешних механических и химических повреждений, воздействия температуры, иссушения, проникновения микробов. Кожа принимает участие в теплорегуляции, газообмене и выведении продуктов распада. Производные кожи могут принимать участие в формировании органов передвижения (копыта), служить для хватания (когти), нападения и защиты (рога, иглы и др.), полёта (складки), плавания (перепонки). Кожа содержит рецепторы органов осязания, в ней много желёз разного назначения (слизистые, жировые, пахучие, потовые и пр.). Кожные покровы позвоночных животных как система наружных органов, которая имеет непосредственную связь с окружающей средой, в первую очередь реагируют на изменения в условиях внешней среды. Поэтому кожа и ее производные у хордовых чрезвычайно разнообразны по строению и функции.
1. Сравнительный обзор кожных покровов у хордовых животных
В индивидуальном развитии первичный кожный покров — слой эктодермы − дает наружный слой кожи, т. е. эпидермис со всеми его производными (пигмент, органы чувств, железы и роговой слой). Собственно кожа, или кориум,— соединительно-тканый слой кожных покровов — развивается за счет мезодермы от кожного листка миотома. Первичным строением кожи можно считать однослойный эпидермис и кориум, состоящий из неоформленной студенистой ткани. Такой кожный покров имеется у ланцетника и у зародышей позвоночных (рис. 1). У асцидий и сальп − кожные покровы срастаются со стенкой тела в общий слой — мантию, а на поверхности тела выделяется особый, содержащий клетчатку покров — туника. Уже начиная с круглоротых и у всех первичноводных позвоночных (бесчелюстных) и у всех рыб (и даже частично у земноводных), строение кожи характеризуется следующими общими чертами:
1) эпидермис многослойный с большим количеством разнообразных слизистых и зернистых железистых клеток, функционирующих как одноклеточные железы;
2) кориум с плотными рядами коллагеновых и эластических волокон, располагающихся правильными чередующимися слоями — продольным и вертикальным.
У современных круглоротых (рис. 1) кожа голая и имеет много слизистых железистых клеток. Она лишена каких-либо костных образований и только в ротовой воронке имеет роговые зубы. У ископаемых бесчелюстных − щитковых имелся панцирь из кожных костей, развившийся у них в связи с донным образом жизни.
У рыб кожные покровы характеризуются наличием различных костных чешуй. Наиболее древними (первичными) являются плакоидные чешуи акуловых рыб. Плакоидная чешуя состоит из костной пластинки с полостью пульпы внутри и зубом из дентина, одетым эмалью, что указывает на генетическую связь с зубами всех позвоночных. Это свидетельство общего происхождения всех позвоночных от акулоподобных предков с плакоидными чешуями. Изучение шлифов чешуи ископаемых рыб, а так же эмбрионального развития чешуи современных костных рыб показывает, что зачатки плакоидных чешуи являются составной частью также и сложных ганоидных и космоидных чешуи.
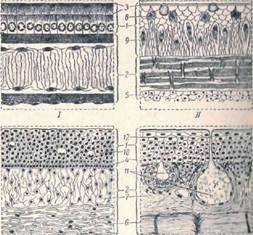
Ш IV
Рис. 1. Разрез через кожу хордовых (по Нилу и Рэнду).
I — ланцетник; II — минога; III — акула IV — лягушка:
I — эпидермис, 2 кориум. 3 — кутикула, 4 − ростковый – мальпигиев слой, 5 − плдкожная соединительная ткань, 6 — плотная и 7 — рыхлая соединительная ткань, 8 — зернистые клетки, 9 — колбовидные клетки, 10 − слизистые клетки, 11 слизистые железы, I2 − проток железы.
В эволюции чешуи костных рыб (рис. 2) можно отметить 3 этапа:
1) усложнение костных чешуи путем срастания отдельных плакоидных чешуи и преобразования дентина в космин — космоидные чешуи кистеперых рыб, ископаемых двоякодышащих, а также Acanthodii из панцирных рыб;
2) дальнейшее усложнение путем развития нового поверхностного слоя ганоина и глубокого слоя изопедина при преобразовании космоидной чешуи в ганоидную у современных костных ганоидов (Holostei), многоперых (Polypterus) и у ископаемых палеонисцид (Paleoniscoidea);
3) позднейшее упрощение костной чешуи путем редукции всех слоев, за исключением никого костного слоя у циклоидных и ктеноидных чешуи, как это имеет место у всех костистых рыб (Teleostei), современных двоякодышащих (Dipnoi) и ильной рыбы (Amia из Holostei).
Кожа современных амфибий имеет еще признаки водных позвоночных. Эпидермис, многослойный, слизистый, кориум с правильным расположением коллагеновых волокон. Однако в коже амфибий вместо разбросанных железистых клеток имеются большие мешковидные сложные железы, погруженные глубоко в кожу (рис.1 (IV)). У современных хвостатых (Caudata) и бесхвостых (Ecaudata) амфибий кожа голая; только у безногих червяг (Apoda) имеются костные чешуйки, погруженные в особые мешочки и коже. У ископаемых амфибий стегоцефалов (Stegocephalia) имелся сильно развитый костный панцирь кожного происхождения, образованный сросшимися костными чешуями. Кожа амфибий является важным органом дыхания и водного обмена. Она лишена роговых образований и богата слизеотделительными железами (сходна с кожей рыб). Но отличается тем, что железы многоклеточные и кожный секрет отсутствует. Для кожи характерно прикрепление к телу не на всем протяжении, а лишь в определенных участках, между которыми располагаются обширные пространства, заполненные лимфой − лимфатические мешки.
Пресмыкающиеся (рептилии)
Кожа имеет покров в виде роговых чешуй и щитков.
Пальцы вооружены развитыми когтями. По внутреннему краю бедер (ящериц) тянется ряд чешуек с отверстиями – бедренные поры. Из которых весной выступают нитевидные образования. Бедренные поры связаны с половой деятельностью. Кожа плотно прилегает к телу. Тело крокодила покрыто роговыми щиткам и бляшками, под которыми располагаются костные щитки (отсюда другое название – панцирные гады).
Особый интерес представляет наружный костный панцирь черепах, развившийся из костного кожного панциря путем срастания его с внутренним скелетом. Черепахи у них пассивный орган защиты – костный панцирь, в котором заключено туловище. Верхний щит этого панциря, носящий название карапакса, образован костными пластинками кожного происхождения, с которыми обычно сливаются расширенные остистые отростки позвонков и ребра. Нижний щит, или пластрон. Как карапакс, так и пластрон покрыты сверху роговыми щитками (у одной группы – мягкой кожей).
Кожные покровы рептилий, птиц и млекопитающих, несмотря на резкое различие по внешним признакам (чешуи, мер им, волосы), характеризуются общими чертами строения, что является важным признаком для объединения всех трех классов высших животных в группу амниот.
Слизистый эпидермис низших позвоночных замещается сухим роговым покровом, предохраняющим кожу амниот от высыхания. Это преобразование эпидермиса для первичных амниот явилось крайне важным приспособлением в процессе их эволюции. Позволяющим им расселиться по суше, так как первичные наземные позвоночные − амфибии были вынуждены жить во влажных местах по берегам пресноводных водоемов.
Таким образом, превращение слизистого покрова в роговой явилось одним из важных факторов биологического прогресса, обусловившим расцвет рептилий в мезозойскую эру, а также расцвет птиц и млекопитающих, заменивших рептилий, в кайнозойскую эру. Ороговение происходит благодаря появлению в плазме эпидермиса зернышек рогового вещества — кератогиалина, элеидина, которые постепенно заполняют всю клетку, после чего она отмирает. Поэтому эпидермис делится на мальпигиев, или ростковый, слой с живыми клетками и роговой слой с отмирающими клетками. Кориум амниот состоит из сложного переплетения соединительнотканых волокон (коллагеновых и эластических). Кожа рептилий, вследствие ороговения эпидермиса, лишена желез; остаются только железы специального назначения (бедренные поры яшериц, мускусные железы крокодилов) для всех рептилий характерно развитие роговых чешуй разнообразного строения: простые бугорки, чешуи, плоские пластинки, стилевидные выросты. У некоторых пресмыкающихся (крокодилы, хамелеоны, гекконы) под роговой чешуей сохраняются костные пластинки. Изучение развития чешуи рептилий, а также изучение покровов ископаемых амфибий и рептилий позволяет сделать заключение о происхождении роговой чешуи рептилий из костной чешуи предков земноводных путем прогрессивного развития рогового слоя эпидермиса и редукции костной пластинки (рис. 2). Кожа птиц очень близка по строению к коже рептилий; она также лишена желез (кроме копчиковой железы над хвостом), но значительно тоньше и подвижнее. Типичные роговые чешуи у птиц имеются лишь на ногах, на теле же развиваются перья, имеющие разнообразное строение и окраску. Строение чешуевидных перьев на крыльях пингвинов (вторично упрощенных в связи с приспособлением к нырянию под водой) и на ногах страусов, а также на голове археоптерикса позволяет сделать заключение о происхождении перьев из чешуевидных образований.
Это вполне подтверждается эмбриональным развитием (рис. 2). Эмбриональный зачаток пера гомологичен зачатку роговой чешуи. Позже он превращается в пальцевидный вырост, эпидермальный чехлик которого на определенной стадии развития распадается на отдельные продольные стержни — бородки. Контурные перья развиваются сложнее благодаря неравномерному росту бородок. Из других специфических производных кожи у птиц имеются: роговой клюв (различного строения в зависимости от характера пищи), когти на пальцах ног, а у некоторых птиц и на I пальце крыла (у археоптерикса на грех пальцах крыла) и, наконец, различные кожные выросты — гребни, бородки, сережки и т. д., играющие роль вторичных половых признаков.
1а-1д — ряд стадий дентинообразовання у акуловых рыб при развитии плакоидных чешуи с типичным эмалевым органом, 2а — 2г развитие костных зубчиков (2а~2б), спинных жучек и бляшек (2в—2г) в коже осетровых рыб с выпадением дентинообразоаания. За—Зв— развитие циклоидных чешуи в коже костистых рыб при полной рудиментации эмалевого органа, 4 — схема ганоидной чешуи как пример комплекса многих генераций чешуи, 5 и 6 — замещение скелетообразующей функции кожи железистой функцией в коже рыб. 7—10 — общий зачаток роговых чешуи, перьев и волос у амниот. 7а— 7г — стадии развития роговых чешуи у рептилий и птиц, 8а —8г — развитие пухового пера и 8д развитие контурного пера, 9а—9г — стадии развития полоса при полной рудиментации соединительнотканого сосочка и прогрессивного развития эпидермального зачатка, 10а—10б − восстановление роговых чешуи в коже млекопитающих
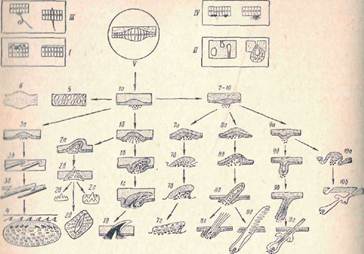
Рис. 2. Схема филогенетического развития чешуи, перьев и волос по данным эмбрионального развития (по Матвееву). / — V — типы функций кожных покровов (/ -защитная, путем образования ресничек и кутикулы; // секреторная, путем образования одноклеточных и многоклеточных желез, /// чувствительность, путем обособления первичных и вторичных чувствующих клеток; IV — окраска, путем дифференциации пигментных клеток; V — защитная функция кожи позвоночных животных, путем развития из общего зачатка):
Кожа млекопитающих в отличие от других амниот богата различными кожными железами: потовыми, сальными, млечными, а также различными железами специального назначения, например: копытные, мускусные, пахучие, околоанальные и т. д. Все эти железы являются производными эпидермиса и лишь вторично глубоко погружены в кориум. Наличие желез в коже млекопитающих при отсутствии их у рептилий и птиц показывает, что предки млекопитающих произошли в процессе эволюции от очень древних рептилий, еще сохранивших общие признаки с ископаемыми земноводными. Млечные железы — типичные кожные железы, происшедшие от простых мешковидных трубчатых желез типа потовых путем их сложного ветвления.
Совершенно своеобразными образованиями кожных покровов млекопитающих являются волосы. В отличие от роговых чешуи и перьев, волосы — чисто эпидермальные образования, и только в луковице волоса имеется соединительнотканый сосочек. Присутствие роговых чешуи у некоторых млекопитающих (неполнозубые, выхухоль, бобр, нутрия и т. д.) одновременно с волосами показывает, что волосы развились самостоятельно между роговыми чешуями, лишь позднее заменив их. Поэтому вопрос о происхождении волос менее ясен, чем эволюция роговых чешуи и перьев. Эмбриональное развитие волос (рис. 2) дает возможность установить гомологию первичного зачатка волоса с зачатком чешуи. Это показывает, что волосы произошли из чешуевидных органов путем их изменения на ранних стадиях развития.
Помимо волос, производными кожи млекопитающих являются различные другие роговые образования: когти, ногти, копыта и, наконец, рога полорогих.
2. Особенности строения кожных покровов и их производных с учетом различных условий жизни хордовых животных
2.1 Особенности строения и образования чешуи
Чешуя рыбы всегда является производным собственно кожи (кориума), и только иногда, кроме кориума, в ее образовании принимает второстепенное участие и эпидермис. Различают четыре основных типа рыбьей чешуи: плакоидную, космоидную, ганоидную и костную.
Плакоидная чешуя − дает начало не только ганоидной и костной чешуе, но и зубам. У акулы, зубы представляют настоящие плакоидные чешуи. Эти зубы вполне гомологичны зубам всех вышестоящих классов, вплоть до млекопитающих, у которых зубы тоже состоят из дентина, покрыты веществом эктодермического происхождения − эмалью и содержат внутреннюю полость, заполненную мякотью.
Космоидная чешуя − особый вид чешуи, которая встречается у некоторых ископаемых костных рыб и встречается у современной латимерии. Она лишена ганоина и поверхностный слой ее состоит из космина, который по своему строению состоит из многих сросшихся друг с другом отдельных дентиновых зубов. Ганоидная чешуя свойственна только очень немногим современным рыбам (многоперовым и каймановым), но зато у ископаемых рыб она имела очень широкое распространение. В типичном случае ганоидные чешуи имеют вид плоских ромбических пластинок, которые расположёны косыми рядами и соединены друг с другом при помощи особых сочленений, так что образуется сплошной панцирь, покрывающий все тело животного. Наружный слой ганоидной чешуи состоит из особого очень твердого вещества — ганоина, нижний из костной ткани. Образуется ганоидная чешуя в соединительной ткани и, следовательно, никогда не бывает покрыта эмалью. Нижний костный слой ганоидной чешуи, по-видимому, образуется из дентина, в который проникают костные клетки. В противоположность плакоидной чешуе ганоидная не сменяется, образуясь на всю жизнь. Эволюция чешуи ископаемых рыб с несомненностью доказывает, что ганоидная чешуя возникла путем срастания основных пластинок отдельных плакоидных чешуи с подслаивающими их костными пластинками. Сверху эти чешуи покрываются ганоином.
Костная чешуя свойственна всем современным костным рыбам, за исключением многоперых, латимерии и каймановой рыбы. Костные чешуи представляют костные пластинки различной величины, черепицеобразно накладывающиеся друг на друга своими краями. Они постоянно растут, образуя годичные кольца по периферии пластинки (рис.3).
Ихтиологи по этим годичным кольцам определяют возраст рыб, Для окуневых характерны ктеноидные чешуи с шипиками по заднему краю чешуи, для карповых, лососевых характерны гладкие циклоидные без зубчиков. У многих донных рыб (сомы, угри) чешуи полностью редуцируются. У осетровых имеются особые костные чешуи, образующие пять продольных рядов жучек с мелкими звездчатыми чешуйками между ними. У многих рыб на плавниках костные чешуи образуют костные шипы.
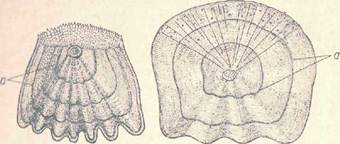
Рис. 3. Чешуя костных рыб. / — ктенондная (окуня); // циклоидная (карповой рыбы): а− годовые кольца
Окраска рыб зависит от ряда причин. Например, серебристый блеск, свойственный не только чешуе, по и многим внутренним органам рыб (плавательному пузырю, брюшине), обусловливается присутствием гуанина.
Гуанин чешуи некоторых рыб (уклейки) используется для технических целей (например, для изготовления искусственного жемчуга). Окраска рыб обусловлена присутствием – хроматофоров. Под влиянием нервного раздражения, могут сжиматься и расширяться, чем обусловлена способность многих рыб менять свою окраску под цвет окружающего фона.
2.2 Особенности строения и образования пера
Для птиц характерна тонкая кожа, полное отсутствие в ней каких-либо костных образований, своеобразный роговой покров, состоящий из перьев. На верхней и нижней челюстях производные кожи — характерные для птиц роговые чехлы, на цевке и пальцах — роговые чешуи, а на концах пальцев — когти. Перья покрывают тело птиц (на примере голубь) не сплошь, а располагаются лишь на определенных участках кожи — птерилиях, между которыми находятся участки, лишенные перьев — аптерии (рис. 4). Такое расположение перьев связано с полетом, так как расположение перьев участками представляет удобство для сокращения мышц во время полета. Лишь у очень немногих, преимущественно нелетающих, птиц аптерии отсутствуют, и перья равномерно покрывают все тело.
Перо (рис. 4) состоит из упругого ствола и более мягких боковых пластинок – наружного и внутреннего опахал. Верхняя часть ствола, к которому прикрепляются опахала, носит название стержня и имеет в поперечном сечении четырехугольную форму, причем верхняя поверхность ствола выпуклая, нижняя же несет продольную борозду. Нижняя, лишенная опахал часть ствола называется очином и имеет в поперечном сечении круглую форму, а у основания снабжена отверстием. В то время как внутренняя часть ствола занята ячеистой сердцевиной, полость очина содержит цепочку из нежных вставленных друг в друга роговых колпачков — душку пера, которая представляет собой омертвевший сосочек, питавший кровью молодое, растущее перо.

Рис. 4. Контурное перо. Очин вскрыт, чтобы показать дужку пера: 1-стержень, 2 – наружное опахало, 3 –внутреннее опахало, 4 – ствол, 5 – очин, 6 – отверстие очина, 7 – дужка пера.
Каждое опахало образовано многочисленными бородками I порядка, на которых сидят более мелкие бородки II порядка. Каждое из них удлиненно-треугольными пластинками — на конце снабжено с крючочками, которые сцепляются с такими же крючочками соседних бородок II порядка, принадлежащих уже другой бородке 1 порядка (рис. 5).
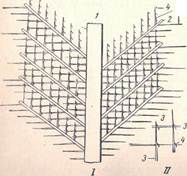
Рис. 5. Схема строения опахала пера (/) и отдельные увеличенные бородки второго порядка (//) (по Гессе) 1- стержень, 2 –бородки первого порядка 3- бородки второго порядка, 4-крючочки
Контурные перья (рис.4) составляют большую часть оперения взрослой птицы, они определяют форму тела животного. Отдельные группы их носят специальные названия: надхвостные, кроющие уха, верхние кроющие крыла. Большие перья, растущие по заднему краю крыла и играющие исключительно важную роль при полете, называются маховыми перьями или махами. Различают маховые 1 порядка, или большие маховые, которые сидят на кисти, и маховые « порядка, или малые, которые прикрепляются к предплечью. Небольшие перья, но имеющие типичное строение маховых и располагающиеся группой на зачаточном I пальце, называются крылышком. Большие перья хвоста, играющие роль руля при полете, носят название рулевых перьев, или рулей. Пуховые перья обычно сидят под контурными перьями, отличаются тонким стержнем и лишены крючочков, благодаря чему не имеют цельных опахал, которые как бы рассучены. Пух представляет собой пуховое перо, стержень которого укорочен, так что все бородки отходят от его вершины одним пучком.
Нитевидные перья и щетинки располагаются у большинства птиц около углов рта и являются пуховыми перьями, сохранившими стержень, но утратившими все бородки; они не имеют никакого отношения к волоскам млекопитающих, несмотря на внешнее сходство с ними.
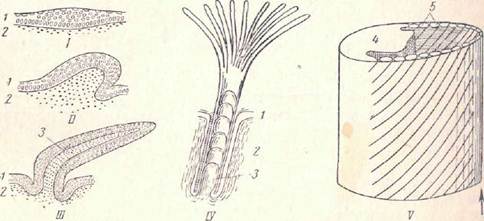
Рис. 6. Схема развития пера (по Гессе). /, // и /// —продольные разрезы через зачатки пера разного возраста; IV —эмбриональное перо в разрезе; V — стереограмма развивающегося контурного пера:
/ эпидермис, 2 — кутис. 3 — сосочек пера, 4 — зачаток стержня, 5 — Зачатки бородок, после сбрасывания наружной кожицы бородки опахала освобождаются и раздвигаются в обе стороны по линии, указанной стрелкой
Расположение маховых перьев приспособлено к полету. Маховые перья имеют сравнительно узкие наружные и широкие внутренние опахала, располагаются в развернутом крыле так, что наружное опахало прикрывает сверху лишь край внутреннего опахала соседнего махового пера. Благодаря такому расположению и тому, что каждое перо способно несколько вращаться вокруг своей оси, при поднимании крыла воздух свободно проходит между перьями; наоборот, при опускании крыла перья образуют сплошную поверхность, оказывающую сильное сопротивление воздуху. Значение этого явления для полета ясно; следует только добавить, что время, которое идет на поднятие крыла значительно меньше времени, которое идет не его опускание. 3начение перьевого покрова. Это очень легкий, прочный и плохо проводящий воздух и тепло покрои, что особенно важно для птиц как для летающих существ, обладающих постоянной и притом очень высокой температурой тела. Кроме того, он придает телу птицы обтекаемую поверхность, а маховые и рулевые перья составляют существеннейшую часть летательного аппарата. Развитие пера протекает следующим образом (рис. 6). Сперва в поверхностном слое собственно кожи образуется скопление мезотермических клеток, которые приподнимают эпидермис в виде бугорка. На этой стадии развития перо сходно с зачатком чешуи пресмыкающегося. Затем этот бугорок разрастается назад, а основание его несколько углубляется в кожу, давая начало влагалищу пера. В дальнейшем соединительнотканная часть бугорка превращается в богатый кровью сосочек пера, а разросшийся эпидермический слой дифференцируется на продольные утолщения, которые лежат ближе к центру, и на тонкий поверхностный слой, образующий чехлик растущего пера. На следующих стадиях продольные утолщения ороговевают и распадаются на бородки эмбрионального пера, которые сбрасывают с себя чехлик. Окончательное (дефинитивное) перо образуется под эмбриональным на том же сосочке, впоследствии сталкивает своего предшественника. Развитие дефинитивного пера протекает в общем сходно с развитием эмбрионального, но одно продольное утолщение (будущий стержень пера) разрастается особенно сильно, а другие утолщения (будущие бородки I порядка) отходят уже от него, так что после спадения чехлика они оказываются расположенными на стержне. Сезонная смена покровов у птиц принципиально ничем не отличается от таковой у рептилий (и млекопитающих). У птиц сменяются все наружные слои покровов и на птерилиях и на аптериях. Окраска перьев зависит от пигмента и от микроскопической структуры пера. Основные пигменты птиц распадаются на две группы: имеющие вид зернышек и палочек меланины и находящиеся в растворе липохромы (близкие к каротиноидам). Меланины обусловливают черный, бурый, серый цвета; липохромы — красный, желтый, зеленый. Черный пигмент в комбинации с желтым обусловливает различные оттенки зеленого цвета, а вместе с красным — различные оттенки красных, коричневых и бурых цветов; желтый пигмент вместе с красным вызывает оранжевую окраску и т. д. Синий и фиолетовый пигменты у птиц неизвестны, и эти часто встречающиеся цвета обусловлены соединением пигментации со сложной микроскопической структурой пера, содержащего многоугольные призматические клетки, преломляющие свет. Но у перьев многих птиц бывает и другого рода окраска, которая непостоянна и изменяется в зависимости от положения птицы по отношению как к источнику света, так и к наблюдателю. Такая металлическая блестящая окраска, отливающая различными цветами, обусловлена отражением световых лучей или от гладкой поверхности пера, или от тончайших пластинок, расположенных на его верхней стороне. Примерами такой окраски может служить оперение скворца, грача, а также перья на зобу и шее голубя.
2.3 Особенности строения шерсти (волосы) и другие образования кожи
Кожа млекопитающих характеризуется толщиной, наличием сосочков на границе между эпидермисом и кориумом, присутствием рыхлой подкожной клетчатки, обилием кожных желез и богатством различного рода роговых образований, из которых главнейшими являются волосы и их видоизменения. Волосы относятся к наиболее характерным роговым образованиям млекопитающих. Они встречаются у представителей только этого класса и свойственны почти всем зверям. Лишь у очень немногих млекопитающих из китообразных (белуха, нарвал) волосы отсутствуют совершенно, у всех же прочих зверей, в том числе большинства китообразных, имеется хотя бы незначительное количество волос (у зародышей китов отдельные волосы имеются на губах). Развитие волоса на первых стадиях протекает сходно с первыми стадиями развития чешуи и пера, т.е. образуется скопление эпидермических клеток. Затем клетки опускаются в глубоко в собственно кожу и дальнейшее развитие волоса протекает своеобразно.
Волос (рис. 7) состоит из двух основных частей: из выдающегося над кожей ствола и из погруженного в кожу корня. В свою очередь ствол волоса состоит из рыхлого сердцевинного вещества, окружающего его плотного коркового слоя и тонкой наружной кожицы. Сердцевинное вещество заполняет так называемый центральный канал волоса и состоит из рыхлой пористой ткани, между клетками которой располагается воздух. Сердцевина имеется у всех диких млекопитающих, но степень развития у разных зверей весьма различна. Сердцевина чрезвычайно непрочна, зато благодаря обилию воздуха оказывает значительное влияние на теплопроводность меха. В связи с этим свойством сердцевина достигает наибольшей толщины у зверей, живущих в холодном климате, в частности у северного оленя, вследствие чего мех этого зверя, вследствие чего мех этого зверя ломкий, но в то же время очень теплый. Плотный корковый слой придает волосу крепость, растяжимость и упругость. Степень развития этого слоя тоже бывает весьма различной. Наконец, роль кожицы заключается в предохранении лежащих под ней слоев волоса от различных физико-химических воздействий.
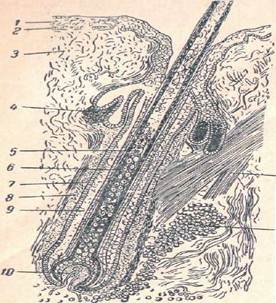
Рис. 7. Продольный разрез волоса в коже (по Паркеру): / — роговой слой Эпидермиса, 2 — его мальпигиев слой, 3 — собственно кожа, 4 — сальная железа, 5 — сердцевина волоса, в — корковое вещество волоса, 7 — стекловидная оболочка между влагалищем волоса и волосяным мешочком, 8 — волосяной мешочек, 9 — влагалище волоса, 10 — сосочек волоса с сосудами, // — жировая ткань, 12 — мышца волосяного мешочка
Снизу ствол волоса постепенно переходит в корень, который заканчивается грушевидным расширением — волосяной луковицей. Последняя, по крайней мере в своей нижней части, состоит из однородных, вполне жизненных клеток. В основание луковицы растущего волоса входит богатый кровеносными сосудами волосяной сосочек, служащий для питания волоса. Корень волоса более или менее глубоко погружен в собственно кожу и сидит в волосяной сумке. Стенки ее образованы двумя слоями: наружным − волосяным мешком, и внутренним, прилегающим к волосу − волосяным влагалищем. Оно является продолжением эпидермиса, который вдается внутрь волосяного мешка и, дойдя до основания волосяной луковицы, заворачивает вверх, прилегая непосредственно к корню волоса. В волосяной мешок обычно открывается проток сальной железы, жировое выделение которой смазывает волос, предохраняя его от смачивания водой. Ниже сальной железы к волосяному мешку прикрепляется пучок гладких мышечных волокон, который другим концом теряется в поверхностных слоях собственно кожи. По сокращении этих мышц волосяной мешок несколько изменяет свое положение, и волос выпрямляется — зверь «ощетинивается». У огромного большинства зверей волосы располагаются в коже не перпендикулярно к ее поверхности, а несколько наклонно. Благодаря этому стержни волос более или менее прилегают к поверхности кожи и обращены вершинам или определенную сторону. Явление это известно под названием ворса. Только немногие роющие звери, например, крот, слепец, которые движутся в своих подземных ходах как вперед головой, так и наоборот, не имеют ворса, так что в каком направлении ни приглаживать их шкурку, волосы всегда ложатся одинаково ровно.
Видоизменением волос являются вибриссы, щетина и иглы; последние играют роль защитного приспособления и особенно развиты у ежен, дикобразов и ехидн. Вибриссы, играющие роль, как уже указывалось, добавочных органов осязания, у многих зверей располагаются, кроме головы, на лапах и на груди, а у крота − на конце хвоста − словом, вообще на местах, которые у того или иного зверя особенно часто соприкасаются с плотной наружной средой.
Мех. Лишь у немногих зверей − у слонов, сирен, носорогов, волосы расположены настолько редко, что говорить о мехе не приходится. Густота и длина меха бывают весьма различными и связаны не только с систематическим положением зверя, но и с его географическим распространением и временем года: как общее правило: звери, живущие в холодном климате. Волосы, составляющие мех, неодинаковы. Различают более длинные редкие волосы, дающие ворс — шерсть, или ость, и густо расположенные тонкие волосы подшерсток. Особенно резко выражена двухярусность и меха, т. е. разделение волосяного покрова на ось и подшерсток, у некоторых водных и полуводных млекопитающих: бобра, выхухоли, утконоса, выдры, морского котика, а также у северных пушных зверей. У некоторых зверей мех состоит преимущественно из подшерстка; таковы крот, слепыш, у других — преимущественно или даже исключительно из ости, например взрослых тюленей, летний мех оленей и кабанов. Линька − процесс выпадения старых волос и замена их новыми может протекать очень постепенно, растягиваясь на длительный срок, и или наоборот, бурно, в коротким срок. Как правило, бурная линька имеет место у зверей обитающих в умеренном и холодном поясах. У большинства зверей в течение года происходят две линьки − весенняя (быстро)и осенняя (медленно).
Собственно кожа (кориум) млекопитающих состоит преимущественно из волокнистой соединительной ткани, отдельные волокна которой образуют сложный переплет. Она всегда богато снабжена кровеносными сосудами, а глубокие слои, состоящие из рыхлой соединительной ткани и носящие название подкожной клетчатки, обычно содержат значительные жировые скопления. Особенного развития достигают жировые скопления водных млекопитающих — китов, тюленей, у которых они образуют толстый слой, сильно уменьшающий теплоотдачу, и у зверей, впадающих в зимнюю спячку: например, сурков, сусликов, летучих мышей, барсуков, медведей. У зверей, впадающих в спячку, жир служит главным образом запасным питательным материалом, и количество его сильно изменяется по временам года: осенью, перед впадением зверя в спячку, он достигает наибольшей толщины; весной же, перед пробуждением, бывает почти весь израсходован.
Эпидермис млекопитающих имеет различную толщину не только у равных групп, но и на разных частях тела одного и того же животного. Наибольшей толщины он достигает на тех местах тела, которые особенно подвержены трению, например, на подошвах ног, ягодицах большинства обезьян (седалищные мозоли), на коленях верблюдов. С поверхности рогового слоя постоянно отпадают отдельные отмершие клетки или группы клеток в виде «перхоти», изнутри же он все время нарастает благодаря делению клеток нижнего, основного слоя эпидермиса, известного под названием мальпигиева. К роговым образованиям наземных млекопитающих относятся ногти, разнообразные когти и копыта (рис. 8). Образования эти совершенно отсутствуют только у китообразных и на большинстве пальцев передних конечностей летучих мышей, а в большей или меньшей степени недоразвиты у сирен и многих тюленей. У всех прочих млекопитающих они развиты хорошо и имеют большое экологическое значение. Доказывается это тем, что не только способ передвижения зверя, но и работа его конечностей связаны с присутствием того или иного из этих образований. Ногти, которые свойственны обезьянам, отчасти и полуобезьянам, имеют более или менее плоскую форму и покрывают конец пальца только сверху. Кроме того, у ногтя хорошо развита покрывающая конец пальца снизу мягкая подушечка пальца, которая у когтя лишь слабо выражена. Когти, имеющие наиболее широкое распространение среди млекопитающих, отличаются от ногтей большей толщиной и изогнутостью верхней роговой ногтевой пластинки, которая охватывает конец пальца с боков и выдается острием за передний край его. У когтя также хорошо выражена нижняя стенка — более рыхлая подошвенная пластинка, недоразвитая в ногте. Наконец, копыто, охватывающее конечную фалангу не только спереди и с боков, но и снизу и образующее нечто вроде толстого рогового башмака, представляет дальнейшее усложнение когтя. В копыте различают роговую стенку, которая соответствует ногтевой пластине, роговую подошву и стрелку. Последняя соответствует подушечке когтя (или ногтя), которая в данном случае ороговевает и вдается снизу острым углом в роговую подошву. Копыта свойственны крупным бегающим или лазающим по скалам млекопитающим.

Рис. 8. Продольный разрез через концы пальцев млекопитающих (по Боасу). / — обезьяна, II — человек, III — хищник, IV — лошадь, 1 — когтевая или ногтевая пластинка , 2 — роговая стенка копыта, 3 — подушечка, 4 — стрелка, 5 — подошвенная пластинка
2.4 Кожные железы у хордовых. Функции, особенности строения
Кожные железы рыб как и у всех позвоночных, являются производным эпидермиса, но, в отличии от вышестоящих классов, эти железы, как и у круглоротых, имеют одноклеточное строение. Наиболее многочисленными являются бокаловидные железы, выделяющие слизь непосредственно наружу. Многочисленные одноклеточные кожные железы, так же как у акулы и миноги, выделяют обильную слизь, которая играет роль смазки, ослабляющей трение тела рыбы о воду при плавании. Кроме того, встречаются еще шаровидные и колбовидные железы, секрет которых выделяется в межклеточные пространства. У некоторых рыб имеются еще особые ядовитые железы, располагающиеся обычно у основания острых плавниковых лучей или у основания шипов, сидящих на заднем краю жаберной крышки. Выделяемое ими белковое вещество представляет сильнейший яд, который даже у человека может вызвать серьезные последствия. Наконец, органы свечения, которые встречаются у многих глубоководных рыб, — тоже железистые образования. Кожа амфибий богата слизеотделительными железами (сходна с кожей рыб). Но отличается тем, что железы многоклеточные и кожный секрет отсутствует. Кожа рептилий, вследствие ороговения эпидермиса, лишена желез; остаются только железы специального назначения − бедренные поры яшериц, мускусные железы у крокодилов. Единственная железа у птиц — копчиковая — расположена над корнем хвоста. Она выделяет секрет, которым птица с помощью клюва смазывает перья. Этот жирный секрет предохраняет перья от смачивания водой. Особенно сильно развита копчиковая железа у водоплавающих птиц, у некоторых же наземных форм отсутствует. Кожные железы млекопитающих бывают двух основных типов: сальные и потовые.
Сальные железы имеют гроздевидное (альвеолярное) строение, так как канал их разветвляется на отдельные канальцы, которые заканчиваются расширениями — альвеолами. Открываются сальные железы почти всегда в волосяной сумке и выделяют кожное сало, которое служит смазкой для кожи и волос, предохраняя их от высыхания и смачивания. У зародышей кожное сало густым слоем покрывает все тело и облегчает прохождение плода при родах через половые пути матери. Потовые железы имеют строение простых трубочек, конечные части которых свернуты клубком. Открываются они либо непосредственно на поверхности кожи, либо в волосяную сумку и выделяют пот. В противоположность кожному салу пот не вырабатывается в клетках железы, а поступает в железу непосредственно из кровеносных сосудов. По химическому составу пот близок к моче и содержит в растворенном состоянии мочевину, поваренную соль и т. п., но воды в нем значительно больше, чем в моче. Хотя при отделении пота из организма выводятся некоторые вредные вещества, но главная роль пота заключается в том, что, испаряясь, он охлаждает поверхность тела. Таким образом, потовые железы служат терморегулирующими органами. Потовые железы отсутствуют у ряда млекопитающих — так, их совершенно нет у китообразных, кротов, ящеров; у грызунов они развиты слабо, а у кролика имеются только на губах и в паховой области. Пахучие железы имеют более сложное строение, чем предыдущие, и чаще представляют видоизмененные потовые железы, реже сальные, но иногда комбинацию и тех и других, и открываются в таком случае в особые мешки. К пахучим железам относятся заднепроходные железы многих куньих, которые выделяют сильно вонючий секрет и, с одной стороны, служат для самозащиты животных (американские скунсы, или вонючки — Mephitis, отчасти наши хорьки), с другой помогают особям разного пола отыскивать друг друга (мускусные железы самцов кабарги, находящиеся в особом мешочке на брюхе, выхухоли и бобра, располагающиеся у корня хвоста, и т. д.). Слезные мешки многих жвачных (оленей, овен, антилоп). Копытные железы козлов и некоторых других млекопитающих, которые служат, по-видимому, не только для смазывания трущихся друг о друга поверхностей копыт, но и для разыскивания особей одного вида, так как они оставляют запах на следу. Млечные железы являются видоизмененными потовыми железами. Однако, исключая однопроходных (утконос и ехидна), они имеют не трубчатое строение, как типичные потовые железы, а гроздевидное. У вcex млекопитающих, исключая однопроходных, млечные железы снабжены coсками и исходном положении или тянутся парными рядами от подмышечной до паховой области. Такое расположение свойственно насекомоядным, хищникам и свинья, насекомоядным, хищным и свиньям, многим грызунам. Иногда из этого парного ряда утрачиваются только средние соски, но чаще либо все передние, главное, что соски сохраняются только в паховой области (большинство копытных, китообразные), либо, наоборот, утрачиваются все задние, так что сохраняется только грудная пара (обезьяны, сирены, слоны, огромное большинство − летучих мышей). Число сосков варьирует от 2 до 14. Как более или менее общее правило, число сосков приблизительно соответствует числу детенышей в помете; на одну пару сосков приходится один детеныш. Различают истинные соски, у которых протоки млечных желез открываются многочисленными отверстиями на конце соска, и ложные — у которых протоки желез открываются на дне более или менее длинного общего канала, прободающего весь и сосок. Первый тип свойствен обезьянам, полуобезьянам и некоторым грызунам, второй — хищным и копытным. Между этими двумя основными типами сосков имеются и переходные.
Молоко по составу и способу отделения сильно отличается от всех прочих секретов, выделяемых организмом. Оно состоит из смеси основных питательных веществ, соединенных в наиболее удобно усвояемом виде, и является незаменимой пищей детеныша. Хотя у разных зверей соотношение веществ, входящих в состав молока, несколько изменяется, оно всегда содержит белок, молочный сахар, жир, небольшое количество минеральных солей и много (около 90%) воды. В противоположность другим железам млечные железы начинают функционировать у самок только на поздних стадиях беременности, и выделение молока происходит под влиянием гормонов, которые вырабатываются, по-видимому, в теле зародыша и попадают в кровь матери.
Заключение
Итак, кожа позвоночных образующая прочный покров, представляет собой очень важный в функциональном отношении орган. Она не только защищает тело от механических повреждений, излишней потери воды, вредных действий сильных световых раздражений и проникновения различных болезнетворных организмов, но принимает более или менее важное участие в процессе обмена веществ, т. е. несет функцию добавочного, иногда весьма существенного, органа дыхания, а также выделения через кожные железы. Кожа позвоночных в противоположность беспозвоночным, состоит всегда из многих слоев клеток, которые образуют два основных слоя: наружный эпидермис, и кориум. Оба слоя кожи отличаются друг от друга и строением и происхождением: кориум имеет волокнистое строение и развивается из мезодермы, а именно из наружного листка миотомов зародыша, тогда как эпидермис имеет эпителиальный характер и развивается из эктодермы.
Оба слоя кожи могут давать начало различного рода скелетным образованиям: кориум − плакоидной чешуе низших рыб и ее производным (зубам, кожным окостенениям и костной чешуе высших рыб), эпидермис — роговой чешуе пресмыкающихся и ее производным (перьям птиц и волосам млекопитающих). Эпидермис, кроме того, дает начало кожным железам.
Список использованной литературы
1. Матвеев Б. С. и др. Курс зоологии, том 2, Москва, Высшая школа, 1966.
2. Наумов С. П. Зоология позвоночных. Москва, Просвещение, 1982.
© 2010 Интернет База Рефератов