
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Курсовая работа: Механизмы выживания бактерий в окружающей среде
Курсовая работа: Механизмы выживания бактерий в окружающей среде
МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РОССИЙСКОЙ ФЕДЕРАЦИИ
Оренбургский государственный аграрный университет
Факультет ветеринарной медицины и биотехнологии
Кафедра микробиологии
КУРСОВАЯ РАБОТА
по микробиологии на тему:
Механизмы выживания бактерий в окружающей среде
Работу выполнил:
студент III курса
специальности «Микробиология»
Акжигитов Абай Сарсенгалиевич
Научный руководитель:
доцент кафедры
Киргизова Светлана Борисовна
Оренбург – 2010
Содержание работы:
Введение
1. Механизмы выживания бактерий при низких температурах
2. Механизмы выживания бактерий при высоких температурах
2.1 Молекулярные механизмы термофилии
2.1.1 Особенность липидов и мембран
2.1.2 Термостабильность макромолекул
3. Механизмы выживания бактерий при экстремальных значениях рН
3.1 Основные механизмы выживания
4. Жизнь бактерий при высоких концентрациях солей, растворенных веществ и в условиях недостатка воды
5. Реакции бактерий на тяжелые металлы и токсичные вещества окружающей среде
6. Жизнь микроорганизмов в условиях интенсивного облучения
6.1 Защитные механизмы
6.1.1 Механизмы репарации ДНК
6.1.2 Механизмы резистентности Micrococcus radiodurans
7. Роль стрессосом как факторов выживания микроорганизмов
Заключение
Список литературы
ВВЕДЕНИЕ
Выяснение причин того, каким образом бактерии адаптируются к различным условиям окружающей среды, имеет очень важное значение для науки. Информация об этих процессах дала бы понимание основных механизмов выживания некоторых из наиболее устойчивых организмов на Земле. Разнообразные группы бактерий могут развиваться в условиях, не доступных, для других организмов.
Огромному разнообразию условий, представляемых биосферой бактериям, соответствует разнообразие их свойств и адаптаций. Обладая огромной численностью популяций и выработанными эволюцией механизмами изменчивости и диффузии генетических детерминаций, большинство бактериальных видов находится в состоянии постоянного адаптационного движения в соответствии с постоянно изменяющимися условиями среды. Бактерии обладают весьма сложными и совершенными механизмами молекулярных адаптаций, о существовании которых еще относительно недавно нельзя было даже предположить.
Если рассматривать жизнь в окружающей среде, то очевидно, что природа может быть весьма враждебной, и что организмам приходится бороться за жизнь. Давно известно, что жизнь встречается в самых суровых условиях: в горячих источниках, которые нередко отличаются повышенной кислотностью; в соленых озерах и солеварнях; в источниках с повышенной кислотностью (например, в рудничных стоках), которые могут содержать также токсичные тяжелые металлы в высоких концентрациях; на сухих поверхностях скал и в пустынях; при температурах около точки замерзания воды и даже ниже ее.
Большой интерес к адаптации микроорганизмов к разнообразным условиям окружающей среды был вызван и долгое время поддерживался поисками жизни на других планетах. Ведь зная границы жизни на Земле, можно составить представление о физических и химических пределах, в которых жизнь могла возникнуть не только на нашей, но и на других планетах.
Изучение механизмов выживания бактерий также имеет очень важное значение для медицины, ветеринарии, фармакологии. Устойчивость микроорганизмов к различным факторам среды ставит вопрос о специальных, новых методах стерилизации и хранения продуктов, что играет важнейшую роль для пищевой промышленности. Таким образом, тема данной курсовой работы является весьма актуальной в настоящее время.
В данной курсовой работе речь будет идти об условиях окружающей среды, в границах которых способны существовать микроорганизмы, а также о механизмах, при помощи которых они выживают в этих условиях.
Цель работы: исследование молекулярных и морфологических механизмов выживания бактерий в различных условиях окружающей среды.
Предмет исследования: бактериальные микроорганизмы и механизмы их выживания.
Основные задачи:
1) изучить механизмы выживания бактерий в условиях низких и высоких температур;
2) рассмотреть механизмы выживания бактерий при экстремальных значениях рН;
3) дать описание механизмов выживания бактерий при высоких концентрациях солей и растворенных веществ;
4) описать защитные реакции бактерий на действие тяжелых металлов и токсичных веществ в окружающей среде;
5) подробно рассмотреть механизмы выживания бактерий в условиях интенсивного облучения;
6) сделать общее заключение по теме курсовой работы.
1. МЕХАНИЗМЫ ВЫЖИВАНИЯ БАКТЕРИЙ ПРИ НИЗКИХ ТЕМПЕРАТУРАХ
Действие температуры на рост микроорганизмов может быть обусловлено ее непосредственным влиянием на скорость химических реакций и на состояние макромолекулярных компонентов клетки (вязкость мембран, конформацию белков и т.д.). В отличие от теплокровных животных микроорганизмы не могут регулировать свою температуру. Их функциональная активность определяется температурой окружающей среды.
Для микроорганизмов, обитающих в холодных условиях, наиболее важно выжить в те периоды, когда физические и химические факторы окружающей среды препятствуют росту и обмену веществ. Микроорганизмы, растущие только в узкой области температур, могут выжить в условиях с весьма неустойчивыми температурными характеристиками лишь в том случае, если они обладают особым механизмом, который не дает им погибнуть при экстремальных температурах; таким механизмом может быть, например, образование спор или цист (Кашнер Д., 1981) .
При минимальной температуре и дальнейшем ее понижении микроорганизмы в основном не погибают и могут длительное время (например, до наступления теплого сезона) сохранять жизнеспособность. При пониженной температуре снижается не только скорость роста, но и скорость отмирания, и соответственно увеличивается выживаемость организмов. Микроорганизмы способны выжить в неустойчивых суровых условиях в состоянии, близком к анабиозу, например в сухих долинах Антарктики и зонах вечной мерзлоты, в течение тысячелетий.
Согласно современным представлениям, некоторые микроорганизмы способны развиваться при низкой температуре благодаря следующим особенностям:
1) клетки содержат ферменты, имеющие низкую температуру активации, и в связи с этим способные наиболее эффективно функционировать при низкой температуре; при температуре выше 30 °С данные ферменты прекращают свою деятельность;
2) проницаемость мембран несмотря на низкую температуру, остается высокой в связи с большим количеством ненасыщенных жирных кислот, содержащихся в липидах, в результате мембраны не замерзают;
3) не утрачивается свойство образовывать полисомы при низкой температуре (Лях С.П., 1976).
Приспособление к пониженной температуре проявляется в изменении состава мембран (в ней повышается содержание ненасыщенных жирных кислот) и синтезе криопротекторов (например, глицерола). Другой механизм связан с накоплением в клетках больших количеств наиболее важных ферментов, так что даже при неоптимальной температуре их функционирование позволяет клетке поддерживать достаточную активность.
Существуют автохтонные популяции микроорганизмов, которые отличаются низким, но постоянным уровнем активности и использованием для питания органических веществ, уже присутствующих в почве и зимогенные популяции, развивающиеся при поступлении в почву свежих соединений, которые они могут использовать. В устойчивых холодных условиях, например, встречаются представители автохтонной и зимогенной популяций. В холодных местообитаниях с неустойчивым температурным режимом, таких, как верхние слои атмосферы или некоторые полярные районы, где температура в разное время года может колебаться от —50 до 5°С, представлена преимущественно зимогенная популяция. В неустойчивых холодных условиях активность микроорганизмов проявляется только во время короткого периода «оттаивания» летом (в зависимости от температуры и доступности воды). По мере понижения температуры организмы должны переходить в такое физиологическое состояние, благодаря которому они остаются жизнеспособными, хотя и покоящимися, в течение всей зимы. Зимогенные бактериальные популяции в местах с постоянной низкой температурой, например в большинстве морских вод и осадков ниже термоклина, сохраняют способность к дыханию даже при температуре, при которой они не проявляют активности по другим показателям. Выживание этих организмов определяется иной совокупностью факторов по сравнению с активной микробной популяцией, существующей в суровых условиях. Автохтонную бактериальную популяцию можно отличить от зимогенной в холодных условиях с устойчивым температурным режимом на основании области температур их роста. В холодных условиях с неустойчивым режимом встречается только зимогенная популяция. Зимогенные бактерии обычно отличаются широкой областью температур роста, поскольку наиболее важным фактором для их выживания является эффективный вторичный метаболизм, который должен осуществляться при температурах ниже оптимума роста.
Хорошо известно, что бурный рост микроорганизмов в природе происходит в ответ на изменение тех или иных физических факторов, таких, как температура и степень влажности в неустойчивых условиях или состав и концентрация питательных веществ в устойчивых условиях. За периодом активного роста следует лаг-фаза, во время которой микроорганизмы осуществляют, по-видимому, процессы вторичного метаболизма, т. е. прекращают делиться и образуют синтетазы, превращающие первичные продукты обмена веществ во вторичные. Как правило, вторичные метаболиты не обладают биологической активностью; однако некоторые из них, например антибиотики и токсины, могут оказывать вторичный эффект, но обычно не на образующие их клетки, а на другие организмы. Длительное выживание бактериальных клеток связано с успешным протеканием вторичного метаболизма. Образование различных морфологических структур, например спор у Bacillus, удлинение «стебелька» у Caulobacter crescentus, образование кокковидных клеток у Arthrobacter и родственных организмов, а также возникновение цист у Azotobacter связаны с вторичным метаболизмом.
Установлено, что микроорганизмы выживают лучше всего, если вторичный метаболизм протекает в оптимальных условиях. Условия, оптимальные для вторичного метаболизма, всегда отличаются от оптимальных условий для роста и первичного метаболизма. В частности, оптимальная температура для вторичного метаболизма нередко бывает на 20°С ниже оптимума роста соответствующего организма и обычно лежит в сравнительно узком интервале 5—10°С. Следовательно, микроорганизмы должны иметь два четких температурных оптимума: один для роста, а другой для вторичного метаболизма; эффективное осуществление последнего обеспечивает выживание.
Микроорганизмы, выделенные из холодных местообитаний с весьма неустойчивым температурным режимом, являются психротрофами. Они способны расти при температурах на 20°С и более превышающих самую теплую температуру in situ. Выживание этих организмов при температурах ниже минимума их роста обусловлено процессами вторичного метаболизма, которые протекают при температуре на несколько градусов ниже температуры роста этих организмов in situ, и разница между температурами роста и замерзания не превышает 3—5°С. Вторичный метаболизм осуществляется перед самым замерзанием, в результате которого клетки переходят к состоянию покоя. Такой переход необходим, так как очевидно, что бактерии в логарифмической фазе роста более чувствительны к резким изменениям окружающих условий по сравнению с клетками в лаг-фазе.
Одно из очевидных преимуществ, которым обладают психротрофные бактерии в холодных условиях как с устойчивым, так и с неустойчивым режимом, заключается в том, что in situ при субоптимальных для этих бактерий температурах процессы первичного метаболизма протекают у них с очень низкой скоростью и, следовательно, более эффективно. Это особенно важно в связи с низким уровнем содержания питательных веществ в местах обитания в большинстве холодных районов, так как чрезмерно быстрый метаболизм привел бы к быстрому истощению имеющегося источника питания, что вызвало бы голодание клеток, а в дальнейшем - их гибель. Поэтому зимогенные бактерии в морях и океанах иногда не проявляют активности в столбе воды или осадках, так как они обычно находятся в морских микроусловиях, для которых характерно постоянное высокое содержание органических питательных веществ. Примерами таких микроусловий могут служить условия, существующие в кишечнике рыб и беспозвоночных, на поверхности микро- и макрофитных растений и животных, в слое воды на поверхности освещенной зоны и некоторых мелководных осадков
Психрофильные микроорганизмы чаще всего находят в морях. Практически все психрофилы представлены грамотрицательными палочковидными формами. Психрофилию грамотрицательных бактерий обычно рассматривают в связи с особыми свойствами их мембран. Это справедливо для тех психрофилов, максимальная температура роста которых равна 10— 15°С. Тем не менее обнаружено множество грамположительных бактерий, которые либо соответствуют определению психрофилов, либо имеют максимальную температуру роста, лишь ненамного превышающую 20°С (Кашнер Д., 1981).
В местах с неустойчивой низкой температурой, например в Антарктике, грамотрицательные бактерии, а также грамположительные аэробные и анаэробные спорообразующие организмы встречаются редко. В районах с устойчивой низкой температурой среди бактерий преобладают грамотрицательные палочковидные формы, главным образом Pseudomonas, Vibrio и другие близкие им организмы, обладающие окислительной активностью.
Представители другого класса бактерий, которые распространены преимущественно в холодных условиях с неустойчивым температурным режимом, включают грамположительные кокки и палочки, относящиеся главным образом к родам Arthrobacter, Corynebacterium, Brevibacterium, Kurthia, Cellulomonas, а также родственные им организмы. Эти бактерии преобладают в антарктических и арктических почвах, озерах, ледниках, льду и снегу, а также в ледяных пещерах и в верхних слоях атмосферы. Эти организмы отличаются тем, что они способны существовать в очень суровых условиях окружающей среды.
Arthrobacter и родственные ему организмы отличаются тем, что они образуют кокковидные клетки во время стационарной фазы роста или в условиях голодания. Некоторые почвенные виды Arthrobacter способны выдерживать длительные периоды голодания in situ, чем и объясняется их широкое распространение в почвах при неустойчивых низких температурах. Один из видов Arthrobacter, обитающий в морях, имеет несколько температурных оптимумов в области роста примерно от 0 до 40°. Вблизи каждого температурного оптимума у этого вида возникают морфологические изменения. Arthrobacter crystallopoietes способен дольше существовать в условиях голодания, чем грамотрицательные бактерии. Длительное выживание этого вида объясняется его способностью к эндогенному метаболизму в отсутствие питательных веществ в окружающей среде.
Обнаружено, что Corynebacterium erythrogenes в отличие от морских психрофилов сохраняет жизнеспособность при температурах, на 3—5°С превышающих максимальную температуру роста.
Различные виды Bacillus сравнительно редко встречаются в полярных районах, так как они не могут прорастать после образования спор. Большинство бактерий этих видов растет при 25—30°С, но способно к спорообразованию при 0°С (Лях С.П., 1976).
2. МЕХАНИЗМЫ ВЫЖИВАНИЯ БАКТЕРИЙ ПРИ ВЫСОКИХ ТЕМПЕРАТУРАХ
Из широкого набора воздействий, которые окружающая среда может оказывать на организмы, к числу наиболее экстремальных, несомненно, относится повышенная температура. С повышением температуры скорость роста микроорганизмов вначале увеличивается, достигая максимальной, но дальнейшее увеличение температуры ведет к необратимой инактивации клеточных компонентов, прежде всего денатурации белков и нуклеиновых кислот, и гибели клетки. Для большинства организмов характерен весьма незначительный интервал между оптимальной и максимальной температурами.
Для ученых представляет большой интерес изучение микроорганизмов, которые не только выживают, но и размножаются, часто облигатно, при температурах, препятствующих в норме существованию каких бы то ни было форм жизни, вследствие разрушения необходимых для них макромолекул.
Термофилия включает в себя множество молекулярных механизмов и не может быть объяснена каким-либо одним свойством организма. Многочисленные сравнительные физико-химические исследования белков термофильных показали различные механизмы выживания бактерий при высоких температурах (Логинова Л.Г., 1977).
2.1 Молекулярные механизмы термофилии
Возможность существования термофилов при высокой температуре обусловлена следующими особенностями:
1) составом липидных компонентов клеточных мембран, а именно высоким содержанием длинноцепочечных С17—С19 насыщенных жирных кислот с разветвленными цепями;
2) высокой термостабильностью белков и ферментов (последние имеют низкую молекулярную массу и содержат значительное количество ионов кальция);
3) термостабильностью клеточных ультраструктур.
Метаболизм термофильных микроорганизмов соответствует их существованию при высоких температурах. В частности, ферменты отличаются не только высокой термостабильностью, но имеют оптимум активности при существенно более высокой температуре, чем их мезофильные аналоги. Несмотря на то что в целом первичная структура этих ферментов различается лишь немного, точечные аминокислотные замены значительно увеличивают термостабильность вторичной структуры. Термостабильность ДНК обеспечивается суперспирализацией, которая достигается действием специфического фермента — обратной гиразы. Высокой термостабильностью отличаются также рибосомальный аппарат и цитоплазматические мембраны, в которых преобладают насыщенные жирные кислоты. В состав мембран гипертермофильных архей вместо жирных кислот входят специфические липиды — углеводородсодержащие бифитаниловые эфиры (Бухарин О.В., 2005).
2.1.1 Особенность липидов и мембран
Липиды термофильных организмов имеют более высокие температуры плавления, чем липиды нетермофильных. Верхний температурный предел роста бактерий определяется температурой плавления клеточных липидов. Возрастание процентного содержания насыщенных и разветвленных жирных кислот при повышении температуры выращивания микроорганизмов приводит к образованию более устойчивой клеточной мембраны. К главным липидным компонентам экстремальных термофилов относятся жирные кислоты, содержащие 17, 18 и 19 атомов углерода.
Липиды играют важную роль в молекулярном механизме термофилии, а их соединения с углеводами (гликолипиды) способствуют термостабильности мембран. Липиды мембран действуют как изоляторы, препятствующие переносу тепла из внешней среды и предотвращающие таким образом тепловую денатурацию растворимых ферментов.
Липидный состав термофилов зависит от температуры среды, и сохранение физического состояния липидов в мембране при изменении температуры обусловлено изменениями ее липидного состава.
Большой интерес представляет термофильный ацидофил Thermoplasma acidophilum. Термофилия этого организма, растущего при температуре 59°С, связана с наличием длинных изопреновых цепей липидов. Большая часть содержащихся в мембране нейтральных липидов этерифицирована жирными кислотами, а основная масса глико- и фосфолипидов состоит из длинных цепей с простыми эфирными связями. Аминокислотный состав мембранного белка характеризуется относительно низким уровнем заряженных аминокислот и довольно высоким содержанием остатков цистеина. Высокое содержание кислых аминокислот, является обязательным условием для его роста в термофильных условиях.
Большое количество данных о свойствах мембран и их регуляции собрано при изучении используемой в качестве модельной системы Escherichia coli. Обнаружено, что мембраны клеток Е. coli во многих отношениях напоминают мембраны термофильных организмов. Об этом говорят следующие особенности мембран E.coli: 1) соотношение насыщенных и ненасыщенных жирных кислот, накапливающихся в результате подавления биосинтеза липидов, сильно зависит от температуры; например, фракция свободных жирных кислот из клеток, выращенных при 15°С, содержит в 10 раз больше ненасыщенных жирных кислот, чем фракция, полученная из клеток, выращенных при 43°С; 2) при повышении температуры липиды мембраны Е. coli обнаруживают фазовый переход из упорядоченного в неупорядоченное состояние; синтез мембран, содержащих менее одной трети нормального количества таких липидов, приводит к гибели клетки.
Термофилы способны контролировать физические свойства цитоплазматической мембраны, изменяя ее состав в ответ на изменение температуры. Однако этот общий механизм, действует также и у мезофила Е. coli. Таким образом, температурные границы оптимального функционирования мембраны, от которых, несомненно, зависит величина кардинальных температур роста организма, определяются характером химических модификаций веществ, входящих в состав мембраны. Как и в случае термостабильных по своей природе белков, найденных у облигатных и кальдоактивных бактерий, тонкий механизм (или механизмы), участвующий в стабилизации мембраны, остается пока неизвестным. Молекулярная основа регуляции фазового перехода липидов относится к одной из наиболее важных, но еще слабоизученных сторон функционирования мембраны.
2.1.2 Термостабильность макромолекул
Выделенные из термофильных организмов макромолекулы обнаруживают большую термостабильность, чем их аналоги из мезофилов. Такая повышенная термостабильность, свойственна всем макромолекулам клетки, включая ферменты главных метаболических путей, ферредоксин, рибосомы и флагеллин.
Большинство белков, выделенных из термофилов, относится к ферментам, которые гораздо более стабильны, чем их аналоги из мезофилов. Классическим примером служит фермент миокиназа, которая сохраняет активность после нагревания при температуре 100°С. У тетрамера авидина в присутствии биотина температура перехода в денатурированное состояние составляет 132°С. У фосфофруктокиназ С. pasteurianum и Е. coli температурный переход происходит приблизительно при 70 и 60°С соответственно. Таким образом, основное различие между белками мезофилов и термофилов заключается в том, что все белки термофилов должны функционировать при температурах развития этих организмов.
Определение температуры максимальной активности ферментов также указывает на их повышенную термостабильность, а потеря всей или большей части активности в результате превышения этой температуры на 10—15°С согласуется с представлением о денатурации белка.
Физико-химическая характеристика белков термофильных микроорганизмов показывает, что они во многом гомологичны соответствующим белкам мезофилов: термофильные белки имеют такое же содержание участков с альфа-спиральной конфигурацией и бета-структурой и такое же число гидрофобных групп, как и аналоги из мезофильных организмов. Но в отличие от белков мезофилов, которые обычно денатурируются при температуре ниже 60°С, термофильные белки при повышении температуры от комнатной до 55 или 60° претерпевают, небольшие конформационные изменения, не сопровождаемые денатурацией.
Термофильные белки чаще всего содержат большое число неполярных боковых цепей, поэтому в результате стабилизации дополнительными гидрофобными взаимодействиями возникает повышенная термостабильность таких участков термофильных белков.
Термофильные белки более устойчивы к индуцируемой нагреванием агрегации, что следует из большей устойчивости к тепловой коагуляции цитоплазматических белков термофильных бактерий по сравнению с аналогичными белками мезофильных бактерий. Более низкая способность белков к агрегации связана с тенденцией к понижению изоэлектрических точек этих белков и повышению устойчивости их молекул к развертыванию, благодаря чему неполярные участки остаются скрытыми в глубине молекул
Термостабильность белков обусловлена также более низким содержанием цистеина и особенно вспомогательных сульфгидрильных групп, которые обычно располагаются на поверхности белка и поэтому легко подвергаются окислению.
Также известно, что ионы кальция могут стабилизировать ферменты не только у термофильных, но и у мезофильных организмов, и абсолютно необходимы для термостабильности. Образование дополнительных центров связывания кальция в случае термолизина и альфа-амилазы, очевидно, имеет отношение к повышенной термостабильности этих белков. Так, термостабильность термолизина при температурах приблизительно от 50 до 80°С повышается в результате стабилизации одного или двух участков ионами Са2+.
Степень стабильности белков также определяется специфическим положением в полипептидной цепи определенных аминокислот, поэтому огромное влияние на термостабильность молекулы белка может оказать замена в ней всего лишь одной аминокислоты в процессе различных мутаций генетического материала бактерии (Логинова Л.Г., 1977).
2.1.3 Небелковые стабилизирующие факторы
Некоторые ферменты, выделенные из термофилов, неустойчивы при оптимальной температуре роста данного организма. Это говорит о присутствии в клетках термофилов дополнительных стабилизирующих факторов. Такими факторами могут служить обычные клеточные компоненты, такие, как кофакторы, субстраты, мембраны и внутриклеточная среда, содержащая сильно заряженные макромолекулы, которые обеспечивают необходимую дополнительную стабилизацию ферментов.
Одним из самых наглядных примеров стабилизации фермента субстратом и модификаторами служит глутаминсинтетаза из В. stearothermophilus Этот фермент быстро теряет активность при 65°С, если в среде отсутствуют глутамат и ионы аммония. Глутаминсинтетаза стабилизируется также модификаторами, действующими по принципу обратной связи, особенно аланином, гистидином и ЦТФ. Для стабилизации внеклеточных гидролаз, например альфа-амилазы и термолизина требуется присутствие кальция. Для термостабильности глюкозо-6-фосфатдегидрогеназы В. coagulans необходимо наличие сильно заряженной окружающей среды.
2.1.4 Высокозаряженные макромолекулы внутри клеток
Термофильные бактерии синтезируют белки, которые обладают присущей им самим термостабильностью. Как отмечалось выше, многие ферменты, выделенные из облигатных или кальдоактивных бактерий, не являются стабильными при оптимальных и (или) максимальных температурах роста, хотя, несомненно, они более термостабильны, чем их мезофильные аналоги. Однако при изучении клеточных стенок факультативного термофила В. coagulans было показано, что отдельные гликолитические ферменты в неочищенных экстрактах этого организма, выращенного при 37 и 55°С, так же лабильны, как и ферменты из мезофильных организмов. Функциональным механизмом термофилии у факультативного термофила В. coagulans служит заряд внутриклеточных макромолекул, что подтверждается наличием у этого организма термолабильной глицеральдегид-3-фосфатдегидрогеназы.
Заряженная внутриклеточная среда также имеет важное значение для выживания облигатных и кальдоактивных термофилов благодаря тому, что она может стабилизировать некоторые ферменты, обнаруживающие более низкую термостабильность при оптимальной или максимальной температуре роста.
Концепция стабилизации белков in vivo посредством заряженных макромолекул принята в качестве правдоподобного объяснения механизма термофилии у В. coagulans по следующим причинам:
1) в неочищенных экстрактах клеток, выращенных при любой из двух указанных температур, присутствуют заряженные макромолекулы, которые, однако, из-за разбавления в условиях in vitro не могут полностью проявить своего защитного действия на ферменты. Тем не менее имитация существующей в условиях in vivo заряженной среды путем добавления к клеточным экстрактам (NH4)2S04 или NaCl приводит к полному предохранению фермента от инактивации;
2) в случае очищенной до кристаллического состояния ГФДГ, т. е. в условиях, когда отсутствует взаимодействие фермента с другими макромолекулами, заряд сам по себе стабилизирует фермент;
3) микробная клетка представляет собой высокоорганизованную систему, содержащую лишь незначительное количество свободной внутриклеточной воды, таким образом высокозаряженные макромолекулы играют важную роль в механизме термостабилизации белков в интактной клетке. В условиях in vivo заряд макромолекул достаточен для стабилизации относительно термолабильных ферментов облигатных и кальдоактивных бактерий (Кашнер Д.,1981)
3. МЕХАНИЗМЫ ВЫЖИВАНИЯ БАКТЕРИЙ ПРИ ЭКСТРЕМАЛЬНЫХ ЗНАЧЕНИЯХ РН
Давно установлено, что концентрация водородных ионов играет роль фактора, определяющего границы существования живой матери, и является одним из важнейших факторов, влияющих на рост и размножение микроорганизмов. Концентрация водородных ионов воздействует на ионное состояние, а следовательно, и на доступность для организма многих метаболитов и неорганических ионов: с одной стороны, концентрация ионов водорода непосредственно влияет на клетку, ее электрический заряд, состояние мембраны, возможность протекания окислительно-восстановительных реакций, с другой стороны, — косвенно, определяя ионное состояние металлов, кислот, их доступность и токсичность. Невозможно переоценить ее влияние на стабильность и функции макромолекул в биологических процессах. Значения реакции среды различных природных вод и растворов, где развиваются микроорганизмы (от рН 1 2 в кислых источниках и рудничных стоках до рН 10 в содовых озерах), покрывают почти весь теоретически возможный диапазон значений рН (0— 14). Большая часть организмов наилучшим образом развивается при концентрации ионов водорода, близкой к рН=7, что характерно для многих природных сред. Очень высокие (кислая реакция) или очень низкие (щелочная реакция) концентрации водородных ионов обычно токсичны для большинства организмов. В общем, предельные концентрации ионов водорода, выше и ниже которых известные в настоящее время организмы прекращают рост и размножение, приблизительно равны рН=1, что отмечено лишь для немногих бактерий и грибов, и рН=11, что наблюдается в случае некоторых водорослей, грибов и бактерий. Большинство организмов живет при рН от 4 до 9, причем их оптимальный рост наблюдается в среде, близкой к нейтральной.
Каждому микроорганизму свойствен определенный интервал кислотности или щелочности среды, в котором возможен его активный рост. В зависимости от положения оптимального рН развития различают ацидофильные, нейтрофильные и алкалифильные микроорганизмы. Большинство естественных местообитаний имеет нейтральный или близкий к нему (слабокислый или слабощелочной) рН. Так, если рН пресной воды равен 7,0, то реакция морской воды скорее слабощелочная (рН 7,5 — 8,0).
В почве рН может быть достаточно низким, до 4,0. В таких местообитаниях развиваются организмы, называемые алкали - или ацидотолерантными и умеренными алкали- или ацидофилами. Тем не менее встречаются как природные, так и антропогенные местообитания, которые можно отнести к экстремальным: экстремально кислые с рН < 4,0 и экстремально щелочные с рН >10,0. Развивающиеся там микробные сообщества представлены соответствующими экстремофильными микроорганизмами. Примером ацидофилов служат молочнокислые, уксуснокислые бактерии, многие грибы (Бухарин О.В., 2005).
Меньше примеров алкалифильных организмов, требующих для своего роста значений рН до 10 и выше (уробактерии и многие цианобактерии). Среди бактерий обнаружено несколько видов, устойчивых к щелочной среде (рН 8,5 и выше). Сюда следует отнести Bacillus pasteurii бактерию, расщепляющую мочевину и хорошо растущую при реакции среды, близкой к рН 11. В. alcalophilus, выделенная из сточной воды, способна расти в диапазоне рН 9—11,5. Выделены и другие бациллы, очень устойчивые к щелочной среде. Цианобактерии могут развиваться в природной среде с рН 7,5—10, некоторые из этих бактерий имеют оптимум рН 10 (Покровский В.И., 1999).
Природные среды, имеющие рН, близкий к обычному нижнему пределу (3—4), встречаются довольно часто. Однако среды с рН более низким, чем 3—4, чрезвычайно редки. Примерами среднекислых сред обитания служат многие озера, некоторые истощенные почвы и кислые болота. Такие естественные среды обеспечивают развитие многих эукариотических водорослей, бактерий, растительных и животных форм. Из этих сред было выделено множество малоизвестных бактерий: Bactoderma, Caulobacter, Microcyclus, Planctomyces и Thiovobium, обнаруженных в сфагновых болотах с рН от 3 до 5. Представители уксуснокислых бактерий растут в пределах рН от 3 до 5. Интересным примером является Acetobacter acidophilum prov. sp., у которого оптимум роста находился при рН 3, нижний предел роста при рН=2,8, а верхний—при рН=4,3.
3.1 Основные механизмы выживания
Концентрация водородных ионов в окружающей среде оказывает целый ряд прямых и косвенных воздействий на метаболизм и стабильность клетки. себе Ионы водорода (Н+) обладают уникальными свойствами, отличающими их от других катионов. Они представляют собой протоны, лишенные электронов. В водных растворах они быстро гидратируются и образуют ион гидроксония Н30+. В кислой среде преобладают ионы гидроксония, которые реально существуют в форме гидратированных ионов, причем степень гидратации зависит от концентрации ионов гидроксония и температуры.
Микроорганизмы, обладающие способностью расти и размножаться при низких или высоких значениях рН, имеют определенные механизмы, обеспечивающие их выживание. Преимуществом данных микроорганизмов является то, что при таких условиях резко ограничивается конкуренция других организмов. Например, ацидофил Thiobacillus thiooxidans может использовать в кислой среде ионы двухвалентного железа, которые при рН выше 5 самоокисляются, вследствие чего этот производящий энергию субстрат становится недоступным для данного организма. При высоких значениях рН алкалофил Bacillus pasteurii специфически нуждается в аммиаке для транспорта и окисления таких субстратов, как глутаминовая кислота, изолейцин, треонин, а также промежуточных продуктов цикла трикарбоновых кислот — ацетата, а-кетоглутарата и малата. В этом случае преимущество существования В. pasteurii при высоких значениях рН заключается в том, что щелочная среда обеспечивает доступность NH3 для организма.
Изменения рН окружающей среды могут вызывать у многих микроорганизмов компенсаторные ферментативные сдвиги. Например, Escherichia coli реагирует на повышение кислотности среды синтезом декарбоксилаз аминокислот. Образующиеся в результате амины приводят к снижению кислотности среды. Повышение щелочности среды стимулирует образование дезаминаз аминокислот, что приводит к снижению рН. Большинство активно метаболизирующих щелочеустойчивых организмов имеет склонность к снижению рН среды в процессе роста. Такая реакция является вторичным механизмом устойчивости микробных клеток к щелочной среде. Пока неизвестно, каков первичный механизм, обеспечивающий стабильность клеток и их рост при высоких значениях рН. Большинство облигатных ацидофилов не использует такого вторичного механизма устойчивости, поскольку их активный рост не приводит к повышению рН. В тех случаях, когда рост ацидофилов начинается в среде, близкой к нейтральной, величина рН быстро снижается. Экстремальные ацидофилы не просто переносят низкие значения рН, но действительно нуждаются в ионах водорода для своего роста и стабильности. Примером подобных микроорганизмов служит Т. acidophilum, которая лизируется при рН выше 5. Следовательно, устойчивость клеток к высокой кислотности или щелочности, вероятно, объясняется их структурными или метаболическими особенностями (Кашнер Д., 1981).
Внутриклеточный рН экстремальных ацидофилов обычно не ниже 4,5. Транспорт ионов и синтез АТФ управляются протонной помпой. Внеклеточные белки экстремальных ацидофилов отличаются высокой стабильностью по отношению к кислотам, в то время как внутриклеточные имеют нейтральный или слабокислый оптимум активности. Вместе с тем нейтральный рН часто бывает токсичным для клеток экстремальных ацидофилов. Возможно, это связано с нарушением целостности цитоплазматической мембраны, для стабильности которой требуются высокие концентрации ионов водорода.
Внутриклеточный рН экстремальных алкалифилов также достаточно высок, однако не превышает 9,5. Белки экстремальных алкалифилов обладают большой алкалистабильностью, а также имеют оптимум активности при щелочных значениях рН. Благодаря этому протеазы, липазы и целлюлазы из алкалифильных микроорганизмов широко используют при производстве детергентов. Транспортные процессы в клетках алкалофилов и движение клеток основаны на градиенте Na+, однако синтез АТФ происходит за счет функционирования протонной помпы.
У облигатных алкалофилов в поддержании цитоплазматического рН, более низкого по сравнению с наружным, ведущая роль принадлежит Na+/Н+- антипортеру, катализирующему движение внутрь клетки протонов в обмен на ионы натрия, в которых эти бактерии нуждаются. В любом случае основными барьерами, обеспечивающими необходимый рН у облигатных ацидо - и алкалофилов, служат клеточная стенка и ЦПМ (Бухарин О.В., 2005).
Жизнедеятельность микроорганизмов часто может приводить к изменению рН среды. Так, к подкислению среды ведут окисление сульфидов до серной кислоты тионовыми бактериями, процессы нитрификации, многие брожения. В некоторых случаях бактерии могут регулировать рН среды за счет изменения метаболизма. Наиболее известным примером является двухфазность брожений, подробно изученная в 30-х годах XX в. В. Н. Шапошниковым на примере ацетонобутилового брожения (Clostridium acetobutilicum). При снижении рН до критического уровня в результате выделения масляной кислоты бактерия переключается на образование нейтральных продуктов, ацетона и бутанола.
К подщелачиванию среды приводят дезаминирование белков и аминокислот аммонификаторами, разложение мочевины уробактериями, а также фотоассимиляция С02 (так, в часы интенсивного фотосинтеза значение рН воды в фотической зоне водоема может возрастать на 1 — 2 единицы). Стабильное значение рН среды в некоторых местообитаниях связано с ее буферностью. Наибольшие масштабы имеет карбонат/бикарбонатная система, обеспечивающая постоянство рН вод Мирового океана (Заварзин Г.А., 2001).
Все организмы, растущие при экстремальных значениях рН, располагают, механизмами для поддержания внутриклеточного рН на уровне, близком к нормальным физиологическим величинам. Такие кислотолабильные молекулы, как АТР и ДНК, не смогли бы существовать, если бы внутриклеточная концентрация водородных ионов была такой же, как и во внешней среде. Однако по отношению к внутриклеточной среде трудно применить классическую концепцию рН. Согласно этой концепции, рН является применяемым на практике показателем концентрации или активности ионов водорода в водном растворе, между тем как внутриклеточное содержимое представляет собой коллоидный, а не истинный водный раствор. Измерение величины внутриклеточного рН не дает информации относительно недиссоциированных протонов, связанных с донорными молекулами.
Внутриклеточный рН имеет определенную ценность, так как он дает представление об общих условиях, существующих внутри клетки.
В поддержании градиентов рН в клетке важную роль играет как природа клеточной стенки и мембраны, так и клеточный метаболизм.
4. ЖИЗНЬ МИКРООРГАНИЗМОВ ПРИ ВЫСОКИХ КОНЦЕНТРАЦИЯХ СОЛЕЙ И РАСТВОРЕННЫХ ВЕЩЕСТВ И В УСЛОВИЯХ НЕДОСТАТКА ВОДЫ
В живых клетках вода служит средой, в которой молекулы разных размеров взаимодействуют между собой. Структура воды, в которой находятся растворенные вещества, контролирует все жизненно важные процессы в клетке: действие ферментов и регуляцию их активности, ассоциацию и диссоциацию органелл, структуру мембран и их функционирование. Небольшие изменения в концентрации растворенных веществ и активности воды могут приводить к значительным физиологическим изменениям, поэтому не удивительно, что многоклеточные организмы выработали специальные физиологические механизмы для поддержания постоянного состава не только жидкостей тела, но и внутриклеточной среды. Например, в крови млекопитающих поддерживается равновесие между ионами натрия и калия с помощью сложного гормонального контроля, действующего на уровне почек и основанного на обмене между кровью и тканями.
Однако микробные клетки должны самостоятельно приспосабливаться к внешней водной среде. В качестве «экстремальных условий» можно рассматривать весьма обычные условия, когда клетки растут в растворах, значительно более разбавленных, чем их внутренняя среда, что имеет место у всех пресноводных микроорганизмов. Животные предохраняют от осмотического лизиса клетки своего тела, поддерживая концентрации веществ, растворенных во внеклеточных жидкостях, в соответствии с их концентрациями внутри клеток. Часто их наружный покров совершенно непроницаем для воды. Большинство микроорганизмов покрыто жесткой клеточной стенкой, предотвращающей их лизис в результате высокого осмотического давления, возникающего внутри этих клеток. У простейших, которые имеют более гибкие стенки, проблемы, связанные с высоким осмотическим давлением, решаются другим путем: вода, поступающая в клетки, собирается в сократительные вакуоли, а затем выделяется из них наружу.
Напротив, клетки, растущие при высоких концентрациях растворенных веществ, по-видимому, не способны поддерживать цитоплазму в более разбавленном состоянии. Это было бы возможно только в том случае, если бы клетки были непроницаемы для воды или непрерывно осуществляли активное выделение растворенных веществ. Хотя внутриклеточная среда микроорганизмов по химическому составу сильно отличается от внешней, не известно ни одного вида, который был бы способен поддерживать внутри клеток общую концентрацию растворенных веществ на более низком уровне, чем в окружающей среде (Кашнер Д., 1981).
Известно, что многие внутриклеточные компоненты микроорганизмов нуждаются в высоких концентрациях Na+ и К+. Белки галофилов содержат много аспартата и глутамата, т.е. они более «кислые», в белках устанавливаются новые гидрофобные взаимодействия, приводящие к более плотной упаковке глобул. На поверхности клеток работает механизм «белкового щита» (S-слои), когда наружу экспонируются СООН-группы аминокислот, удерживающие Na+. Эти же группы формируют «гидратированную» оболочку клеток за счет электростатического ориентирования диполей воды. Галофилы осуществляют активный транспорт ионов из клетки, таким образом поддерживая некоторый «осмостаз». Также клетки иногда заменяют Na+ на К+.
Для удержания воды в цитоплазме в условиях высокой солености у галофильных микроорганизмов существуют разнообразные механизмы. Основным механизмом приспособления к осмотическому состоянию среды служит синтез микроорганизмами осмопротекторов (осмолитов, или совместимых растворителей) — низкомолекулярных органических веществ, концентрация которых в цитоплазме уравновешивает внешнее давление (см. табл.1 )
Таблица 1.
Совместимые растворители (осмолиты)
| Организмы | Совместимый растворитель | Минимальная о„. |
| Бактерии-нефототрофы |
Глицин-бетаин, пролин (у грамположительных), глутамат (у грамотрицательных) |
0,97-0,90 |
| Пресноводные цианобак-терии | Сахароза, трегалоза | 0,98 |
| Цианобактерии соленых озер | Глицин-бетаин | 0,90-0,75 |
|
Галофильные аноксиген-ные фотогрофные бактерии Edothiorhodospira |
Глицин-бетаин, трегалоза, эктоин | 0,90-0,75 |
| Экстремально-галофильные археи | КС1 (закачивается внутрь с обменом на NaCl) | 0,75 |
Их состав зависит от концентрации NaCl в среде и не одинаков у разных микроорганизмов. К осмопротекторам относятся некоторые аминокислоты и их производные (глутаминовая кислота, пролин), сахара (в частности, трегалоза), гетерогликозиды, полиспирты, глицин-бетаин. При пониженной водной активности организм находится в условиях осмотического стресса, что приводит к уменьшению скорости роста и снижению общего количества образуемой биомассы (Покровский В.И., 1999).
Адаптация к солености у экстремально галофильных архей (порядок Halobacteriales) основана на аккумуляции ионов К+. Внутриклеточная концентрация ионов может быть выше в 1000 раз, чем в окружающей среде, т.е. ферменты галобактерий работают в солевом растворе. Подобная же стратегия обнаружена у некоторых эубактерий — Salinibacter ruber и представителей порядка Haloanaerobiales. Помимо концентрации солей повышенное осмотическое давление и низкая активность воды создаются высоким содержанием органических веществ. Приспособленные к таким условиям организмы называют осмофилами — это спироплазмы, размножающиеся в нектаре цветов, мицелиальные грибы и дрожжи, обитающие в варенье, сиропах, сухофруктах.
Для микроорганизмов, развивающихся на суше, большое значение имеет приспособление к сухости и контакту с воздухом. Условия водного стресса и опасность высыхания создаются на поверхности скал, камней, деревьев, различных сооружений, в почве, особенно почве пустынь. Основными механизмами защиты от высыхания служит образование слизистых капсул или переживающих клеток (спор, конидий, цист). Высокую устойчивость на воздухе обнаруживают многие микобактерии с высоким содержанием липидов в клеточной стенке. Типичными компонентами микроценозов, развивающихся на поверхности камня и в почве, являются микрококки, артробактеры, нокардии, проактиномицеты и актиномицеты. В целом грамположительные бактерии актиномицетной линии рассматривают как континентальную ветвь эволюции прокариот, приспособившуюся к жизни в наземных условиях (Бухарин О.В., 2005).
Жизнеспособность бактерий в условиях недостатка воды определяется многими факторами температурой, реакцией среды, составом солевого раствора и т. п. Причем бактерии с мелкими клетками устойчивее, чем с крупными; кокки устойчивее палочек; грамположительные бактерии устойчивее к высушиванию, чем грамотрицательные и тем более микоплазмы. Высокой устойчивостью к высушиванию обладают микобактерии, клеточные стенки которых содержат большое количество липидов. Споры не только бактерий, но и других микроорганизмов хорошо переносят высушивание.
В условиях недостатка воды некоторые микроорганизмы обволакиваются гидрофильными слизистыми капсулами, которые активно поглощают влагу. Бактерии, обитающие на корнях пустынных растений, выделяют такие значительные количества гигроскопической слизи, что обеспечивают водой не только самих себя, но и растения.
Существует предположение о том, что при недостатке воды бактерии используют метаболическую воду, образующуюся в клетке в результате окисления органического вещества кислородом воздуха. Так, из 1 кг глюкозы микроорганизм может получить около 600 г воды по уравнению
С6Н1206 + 602 = 6С02 + 6Н20
Устойчивость к обезвоживанию у разных бактерий неодинакова. Например, численность жизнеспособных клеток Pseudomonas, внесенных в воздушно-сухую почву после выдерживания в течение месяца, снижается в 100 раз. В то же время Azotobacter остается жизнеспособным в почве даже через десятки лет ее хранения в воздушно-сухом состоянии Выживаемость азотобактера обусловлена его цистами. Интересны исследования, показавшие, что водный стресс приводит к возрастанию содержания актиномицетов среди других микроорганизмов, обнаруживаемых в почве. Это связано с большей выживаемостью актиномицетов в почве по сравнению с грибами и бактериями. Следовательно, выживаемость микроорганизма в сухой почве существенно возрастает, если он способен формировать те или иные устойчивые формы. Так, вегетативные клетки Pseudomonas довольно чувствительны к водному стрессу, в то время как цисты азотобактера и споры актиномицетов проявляют значительную устойчивость к нему (Покровский В.И., 1999).
Выживаемость актиномицетов. Водный стресс приводит к возрастанию процентного содержания актиномицетов среди всех жизнеспособных микроорганизмов, обнаруживаемых в природных почвенных пробах. Это обусловлено большей выживаемостью актиномицетов в почве по сравнению с грибами и истинными бактериями. Устойчивость актиномицетов к обезвоживанию обусловлена устойчивостью спор.
Таким образом, ясно, что выживаемость бактерий в почве значительно возрастает, если данный организм образует какие-либо устойчивые формы. Вегетативные клетки псевдомонаса обладают чрезвычайно высокой чувствительностью к обезвоживанию, в то время как цисты азотобактера и споры актиномицетов (Streptomyces) значительно более устойчивы.
Arthrobacter не имеет явно выраженной покоящейся или защитной формы в цикле развития. Здесь играет определенную роль переход из палочковидной в шарообразную форму и обратно, который претерпевает Arthrobacter. Шарообразная форма клеток обладает большей устойчивостью к обезвоживанию, чем палочковидная (Кашнер Д., 1981).
5. РЕАКЦИИ МИКРООРГАНИЗМОВ НА ТЯЖЕЛЫЕ МЕТАЛЛЫ И ТОКСИЧНЫЕ ВЕЩЕСТВА В ОКРУЖАЮЩЕЙ СРЕДЕ
Среди микроорганизмов есть формы, устойчивые к действию общих клеточных и метаболических ядовитых веществ (фенол, окись углерода, сероводород и др.), отдельные виды обладают способностью использовать эти соединения в качестве источников питания. Считают, что устойчивость микроорганизмов к токсичным веществам во многих случаях определяется плазмидами.
В выработке устойчивости бактерий к токсичным веществам участвуют трансмиссивные плазмиды, несущие гены множественной устойчивости — R-факторы (от англ. resistance — устойчивость). R - факторы обусловливают устойчивость микроорганизмов к нескольким (девять и более) группам веществ — солям тяжелых металлов, а также антибиотикам, лекарственным веществам, и др. Гены, которые определяют устойчивость бактерий, могут находиться в транспозонах, способных перемещаться в разные участки хромосомы и на плазмиды. Распространению множественной устойчивости бактерий способствует комбинация трансмиссивной плазмиды с транспозоном.
Влияние на микроорганизмы токсичных веществ в небольших концентрациях, не вызывающих их гибели, рассматривают как один из вариантов стрессовых (от англ. stress напряжение) воздействий. В таких условиях включаются специальные механизмы клеточного метаболизма, которые обеспечивают выживание бактерий (Бухарин О.В., 2005).
Микроорганизмы по-разному реагируют на тяжелые металлы в зависимости от вида микроорганизма и концентрации тяжелых металлов в среде. Это справедливо также для мышьяка и сурьмы. Всем микробам в качестве компонентов питания необходимы те или иные тяжелые металлы, такие, как Со, Си, Fe, Мп и Zn. Некоторые микроорганизмы нуждаются также в Мо, V и Ni. Все эти металлы участвуют в основном в ферментативном катализе и должны присутствовать в питательной среде лишь в очень низких концентрациях, обычно порядка нескольких микрограммов на один литр. Ряд микроорганизмов способен осуществлять активный транспорт некоторых из этих элементов внутрь клетки. Существуют бактерии и грибы, которые вырабатывают специальные хелатобразующие вещества, облегчающие проникновение железа в клетку при нейтральных значениях рН. Это проникновение происходит в результате активного транспорта хелатного железа и распада хелата после его переноса через плазматическую мембрану. Даже токсичный ион арсената может проникнуть в клетку путем активного транспорта, как в случае Saccharomyces cerevisiae.
Любой из металлов, а также мышьяк или сурьма в достаточно высоких концентрациях становятся токсичными для микроорганизмов. Проявления этой токсичности могут быть различными, например изменение морфологии клеток или клеточного метаболизма, бактериостаз или гибель клеток. В некоторых случаях возникают более толерантные к тяжелому металлу, мышьяку или сурьме резистентные штаммы, т. е. такие, для воздействия на которые необходима более высокая концентрация токсичного вещества, чем для воздействия на родительские штаммы. Обычно эта резистентность обусловлена генетическими модификациями, часто связанными с плазмидами, а иногда — с половым фактором или с хромосомами. Причиной повышенной резистентности может быть уменьшение проницаемости клетки для токсичного вещества или его биохимическое обезвреживание. Показано, что исключительная резистентность Scytalidium к меди (выдерживает концентрацию CuS04 до 1 М) обусловлена кислой реакцией среды (рН от 2,0 до 0,3) и неспособностью ионов меди проникать в клетки при таких значениях рН, поскольку при реакции среды, близкой к нейтральной, гриб становится чувствительным к 4 • 10-5 М CuSO4. Одни микробы обезвреживают тяжелые металлы, мышьяк или сурьму, вырабатывая вещества, реагирующие с указанными элементами внутри клетки (например, при метилировании ртути или мышьяка) или вне ее, т. е. делают их недоступными для ассимиляции микробом (например, осаждение арсената или арсенита ионами железа в процессе окисления арсенопирита при участии Thiobacillus ferrooxidans). Другие микроорганизмы нейтрализуют токсичные соединения, превращая их ферментативным путем в менее вредные (примером может служить восстановление HgCl2 до HgO. Физиологическое состояние организма также определяет его чувствительность к интоксикации тяжелыми металлами, мышьяком или сурьмой.
Механизм токсического действия тяжелых металлов, мышьяка и сурьмы зависит от природы соединения и рассматриваемого организма. Одни элементы, такие, как Си, связываются в основном с клеточной поверхностью, где и локализуются вызываемые ими повреждения. Другие элементы, например Hg, проникают внутрь клетки, где связываются с определенными функциональными группами, в частности с SH-группами, инактивируя таким образом жизненно необходимые молекулы, такие, как молекулы ферментов, или откладываются в металлической форме. Существуют также дополнительные механизмы токсического действия тяжелых металлов, мышьяка и сурьмы, обусловленные тем, что последние могут: 1) играть роль антиметаболитов; 2) образовывать стабильные осадки или хелаты с важными метаболитами или катализировать распад таких метаболитов, в результате чего они становятся недоступными для клетки; 3) замещать структурно или электрохимически важные элементы, что приводит к нарушению ферментативной или клеточной функции.
Одни микробы окисляют восстановленные формы тяжелых металлов и соединений мышьяка или сурьмы, в то время как другие восстанавливают окисленные формы этих элементов в больших масштабах. При окислении восстановленных соединений металлов по крайней мере некоторые микроорганизмы могут извлекать полезную энергию и восстанавливающую способность. При восстановлении окисленных соединений металлов ряд микробов осуществляет процесс, который является, по-видимому, своеобразной формой дыхания, характеризующейся тем, что окисленные соединения металлов, мышьяка или сурьмы служат частично или исключительно в качестве конечных акцепторов электронов. Такие реакции окисления и восстановления могут иметь фундаментальное значение в перераспределении этих элементов в среде.
В табл. 2 перечислены минералы, многие из которых ассоциированы с рудами, подвергающиеся воздействию микроорганизмов.
|
Таблица № 2. Некоторые природные минералы, содержащие металлы и подвергающиеся воздействию микроорганизмов
|
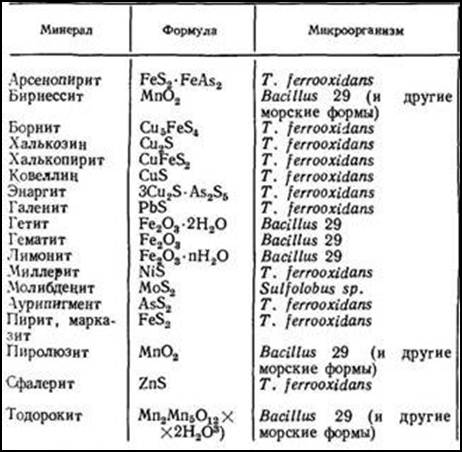
Микробы способны концентрировать тяжелые металлы внутри клеток или на их поверхности. Известны следующие соотношения концентраций различных металлов, содержащихся в морской воде и планктоне: кадмий—1:910, кобальт—1:4600, медь—1:7000, железо — 1 : 87 000, свинец — 1:41 000, марганец — 1 : 9400, титан— 1:20 000 и цинк—1:65 000. В общем конечная концентрация металла внутри клетки может быть на несколько порядков выше его концентрации в окружающей среде. В одних случаях накопление соответствующих соединений оказывается летальным, а в других — нет. На поглощение ионов металлов могут оказывать влияние физиологическое состояние клеток и условия окружающей среды.
Более устойчивы к действию химических веществ из неспорообразующих шаровидные формы. Палочковидные и извитые формы микробов при прочих равных условиях быстрее погибают.
Споры почти не содержат свободной воды, имеют плотную двойную оболочку, поэтому отличаются более высокой устойчивостью к действию химических веществ. Таким образом, действие химических веществ зависит от состава, концентрации, экспозиции, температуры и других факторов (Асонов Н.Р., 1997).
Кислые сточные воды шахт представляют собой пример условий окружающей среды с исключительно высокими концентрациями тяжелых металлов, а также, возможно, мышьяка и сурьмы, токсичными для многих микроорганизмов. Тем не менее, в этих сточных водах была обнаружена смешанная микрофлора, состоящая из водорослей, грибов, простейших и бактерий, которая, по-видимому, специфически адаптировалась к таким условиям.
Изложенное выше показывает, что у некоторых микроорганизмов выработались специфические механизмы взаимодействия с тяжелыми металлами, мышьяком и сурьмой, присутствующими в окружающей среде, иногда в концентрациях, которые токсичны для многих других микробов и высших форм жизни. Микроорганизмы могут использовать эти вещества в качестве источников энергии или акцепторов электронов в процессе дыхания. В ряде случаев у микробов выработались способы удаления этих веществ из среды путем их осаждения, адсорбции или улетучивания. Эти реакции вносят вклад в детоксикацию среды, которая становится более пригодной ие только для микробов, катализирующих такие реакции, но и для других организмов, неспособных развиваться без подобной «помощи» (Кашнер Д., 1981).
6. ЖИЗНЬ МИКРООРГАНИЗМОВ В УСЛОВИЯХ ИНТЕНСИВНОГО ОБЛУЧЕНИЯ
Стимулом для исследования реакции микроорганизмов на облучение послужило стремление решить проблемы, связанные с опасностью радиации для человечества. Хорошо известно, что излучения разных типов обладают потенциальной способностью оказывать на живые организмы разрушительное воздействие.
Однако, если не говорить о высоких дозах, излучения во внешней среде носят такой характер, что для любой клетки существует определенная вероятность избежать повреждения. Исходя из этого, можно было бы предположить, что одноклеточным организмам удается выйти из опасного положения благодаря тому, что они очень быстро размножаются. Тем не менее это, по-видимому, не так, поскольку у них выработались дополнительные средства защиты от летального или повреждающего воздействия облучения. Одноклеточные организмы располагают множеством защитных механизмов, причем многие виды используют не один, а большее число способов борьбы с радиационными повреждениями.
Клеточные механизмы защиты от повреждающего действия радиации настолько широко распространены у микроорганизмов, что трудно оценить опасность, которую представляет для них этот фактор среды. Только тогда, когда защитные механизмы почему-либо не действуют, становятся очевидными реальные размеры опасности, исходящей в основном от коротковолновой части солнечного спектра. Не удивительно поэтому, что начало выяснению природы радиорезистентности было в значительной мере положено обнаружением радиационно-чувствительных мутантов. Такие мутанты не только позволили выявить опасность радиации, но и послужили средствами для исследования механизмов, сдерживающих эту опасность.
Инактивация клеток, вызываемая облучением в относительно низких дозах, обусловлена главным образом повреждением ДНК. Это заключение подтверждается тем, что у прокариот большинство изученных радиационно-чувствительных мутантов дефектны по функциям, имеющим отношение к ДНК. Кроме того, исследование таких мутантов показало, что резистентность обычно достигается не в результате защиты ДНК от индукции повреждении, а скорее благодаря действию механизмов, репарирующих ДНК после того, как повреждения возникли.
Адаптация микроорганизмов к высоким дозам излучений основана на механизмах, участвующих в исправлении повреждений, вызываемых облучением. Известны три независимые системы репарации повреждений ДНК, индуцируемых облучением. Одна из них представляет собой обратную фотохимическую реакцию, происходящую под действием видимого света и фотореактивирующего фермента; вторая — вырезание и замещение поврежденного участка ДНК до ее репликации, а третья — пострепликативную репарацию. Первый из упомянутых механизмов действует только на пиримидиновые димеры, индуцируемые ионизирующим излучением. Многие организмы для защиты от неблагоприятного воздействия радиации используют все три системы. Несмотря на то, что репаративные механизмы имеются у всех исследованных в этом отношении микроорганизмов, последние различаются по своей способности переносить облучение. Самый яркий пример такой вариабельности — исключительно высокая резистентность М. radiodurans и родственных ему видов.
Парадоксально, но одно из последствий облучения — возникновение мутаций — может дать организму преимущество при отборе. Поэтому есть основания предполагать, что с эволюционной точки зрения для организма выгодно установление некоторого равновесия между резистентностью и чувствительностью к радиации; возможно, именно по этой причине защита никогда или почти никогда не бывает полной. Следует ожидать, что соотношение между чувствительностью и резистентностью к облучению неодинаково у разных организмов; и действительно, среди различных видов наблюдается исключительное разнообразие по степени их резистентности к летальному и мутагенному действию облучения. Такое разнообразие создает большие возможности для исследования явлений резистентности и чувствительности организмов к радиации.
Клеточные механизмы, обеспечивающие радиорезистентность, можно разделить на две большие группы. К первой относятся системы, предотвращающие возникновение повреждений в клетке. Вторая включает механизмы, которые восстанавливают (репарируют) повреждения в ДНК, индуцируемые облучением. В настоящем обзоре будут рассмотрены оба аспекта радиорезистентности, но второму из них — репарации — в настоящее время уделяется значительно больше внимания и соответственно он будет обсуждаться здесь более детально. Исследования в этой области были стимулированы одним удивительным открытием: выяснилось, что по крайней мере некоторые из путей репарации более или менее независимы от основных процессов клеточного метаболизма. Поэтому оказалось возможным выделить дефектные штаммы, у которых нарушена система репарации индуцируемых облучением повреждений, но которые тем не менее остаются жизнеспособными. Такие мутанты чувствительны к облучению и в качестве генетических инструментов оказываются исключительно полезными в исследовании клеточных механизмов репарации. Работы в этой области существенно углубили понимание природы радиорезистентности.
Диапазон доз облучения, которому микроорганизмы могут подвергаться эпизодически или постоянно, значительно расширился в последние годы в связи с созданием искусственных источников радиации. В результате этого микроорганизмы стали испытывать воздействие повышенного уровня радиации в окружающей среде. Реакция на такое воздействие представляет как практический, так и научный интерес. Например, при использовании высоких доз облучения для стерилизации пищи возникает проблема, связанная с возможностью индукции или селекции радиорезистентных микроорганизмов, что может иметь неблагоприятные последствия.
Излучение в окружающей среде подразделяется на ионизирующее и неионизирующее. Оба вида опасны для микроорганизмов, но из всех естественных излучений неионизирующая солнечная радиация обладает наибольшим потенциалом биологически вредного воздействия.
Одним из наиболее резистентных к ультрафиолетовому излучению микроорганизмов является Bodo marina, относящийся к морским жгутиковым.
Для инактивации 90% клеток этого организма требуется доза 112 000 эрг-мм-2. Отмечено, что простейшие вообще более резистентны к облучению, чем бактерии (дозы, инактивирующие 90% клеток, колеблются от 5000 до 12 000 эрг-мм-2 для простейших и от 4 до 250 эрг-мм-2 для бактерий). Резистентность разных видов бактерий варьирует в очень широких пределах (Камшилов М.М., 1974).
В отношении резистентности организмов к ионизирующему излучению наблюдается сходная картина. Так, дозы, убивающие 50% клеток в культурах Е. coli, дрожжей, амебы, В. mesentericus и инфузорий возрастают от 5600 до 350 000 Р.
Имеется корреляция между резистентностью организма и уровнем радиации в среде его обитания. Так, микроорганизмы, выделенные из радиоактивных минеральных источников, в 3— 10 раз более резистентны к радиации, чем организмы тех же видов, выделенные из нерадиоактивной воды.
Обнаружен вид Pseadomonas, обитающий в ядерных реакторах, где средняя доза излучения, по-видимому, превышает 106 ФЭР (физический эквивалент рентгена).
В связи с такими случаями явной адаптации микроорганизмов к радиоактивному излучению был поднят вопрос о том, насколько вероятно появление радиорезистентных организмов при использовании облучения для стерилизации. Действительно, Micrococcus radiodurans, обладающий наиболее высокой радиорезистентностью из всех изученных бактерий, был первоначально обнаружен в консервированном мясе, которое подвергалось Y-облучению в дозе несколько мрад. Этот организм может переносить дозы облучения до 500 крад без какой-либо заметной инактивации (Кашнер Д., 1981).
Более устойчивы к излучению грамположительные микробы и менее устойчивы грамотрицательные. Повышенная устойчивость к излучениям отмечена у клостридий ботулизма: они погибают только после воздействия на них дозами в 25—40 кГр. Для достижения стерильности в некоторых случаях необходимо излучение в 50 кГр. Устойчивы к излучению и риккетсии; их устойчивость примерно такая же, как и у спор бацилл.
Некоторые микробы (возбудитель сибирской язвы, кишечная палочка и др.) приобретают устойчивость к излучениям. После нескольких облучений она у них повышается в два или более раза (Асонов Н.Р., 1997).
6.1 Защитные механизмы
Одним из универсальных механизмов адаптации к световому излучению высокой интенсивности и защиты от токсичных форм фотосенсибилизированного кислорода является синтез каротиноидных пигментов. Характерным примером может служить яркая окраска микроорганизмов, живущих в условиях высокой освещенности (в воздухе, на поверхности скал, обнажений горных пород, в высокогорье и т.д.) (Заварзин Г.А., 2001).
Ранние исследования радиационной резистентности были направлены в первую очередь на поиски внутриклеточных веществ, защищающих организм от повреждений. В настоящее время внимание исследователей концентрируется в основном на механизмах, тем или иным способом исправляющих повреждения в ДНК, индуцируемые облучением. Эти механизмы имеют важное значение. Тем не менее представляется вероятным, что определенную вспомогательную роль играют и защитные механизмы.
Для радиорезистентных организмов обычно характерна усиленная пигментация, что чаще всего является причиной резистентности. Пигменты действуют как «энергетические ловушки», препятствующие радиации или ее продуктам достигать ДНК или любых других жизненно важных мишеней.
Резистентность может быть обусловлена также присутствием определенных продуктов метаболизма (внутриклеточных радиопротекторов). Так, Е. coli более резистентна при облучении в присутствии экстрактов из М. radiodurans, чем при облучении в буферном растворе. Защитное действие экстрактов из М. rаdiodurans связано с уменьшением выхода радикалов при участии механизма, аналогичного тому, который действует в отношении известных химических веществ-протекторов.
Своеобразный метаболизм серосодержащих аминокислот у М. radiodurans позволяет думать, что эти аминокислоты выполняют роль сульфгидрильных веществ-протекторов. Зависимость радиорезистентности от концентрации экстракта имеет двухкомпонентный характер: в низких концентрациях он оказывает на тест-организм сенсибилизирующее действие, а в высоких концентрациях — защитное.
Радиорезистентность может определяться уровнем каталазной активности в. клетке. Было показано, что для некоторых бактерий с повышенной радиорезистентностью характерно высокое содержание каталазы.
С возрастанием радиорезистентности увеличивается длина клеток: значительная часть клеток наиболее резистентных штаммов была в 30—40 раз длиннее нормальных. У клеток этих штаммов наблюдалось также своеобразное явление почкования. У резистентных штаммов, упомянутых выше, удлинение клеток было устойчивым признаком, наблюдавшимся в течение трех лет.
К важным факторам, от которых зависит реакция той или иной клеточной системы на любой физический или химический агент, относится состав клеточной стенки. В случае химических мутагенов структура клеточной стенки может определять ее проницаемость, влияя, таким образом, на чувствительность клетки к данному агенту. Хотя структура клеточной стенки не оказывает влияния на проникающую способность ионизирующего излучения, она тем не менее может иметь значение для радиорезистентности микроба. Например, вполне вероятно, что какой-либо связанный с мембраной ферментный комплекс, освобождающийся или активируемый под действием радиации, играет определенную роль в системе (системах) репарации или обусловливает конечную инактивацию клетки. Ионизирующее излучение вызывает освобождение связанной с клеточной поверхностью экзонуклеазы у М. radiodurans. При облучении в дозе 400 крад, сублетальной для этого организма, в клетках остается только 10% фермента, причем степень освобождения фермента зависит от дозы облучения.
Микробы-сапрофиты более устойчивы у световому излучению в сравнении с патогенными. Это объясняется тем, что они чаще подвергаются действию прямых солнечных лучей, поэтому являются более адаптированными (Радчук Н.А., Дунаев Г.В., 1991).
Увеличение содержания ДНК в клетке служит одним из факторов ее радиорезистентности. Это может быть обусловлено либо увеличением числа нуклеоидов в клетке, либо ее полиплоидностью. Нитевидная форма резистентных клеток Е. coli позволяет предполагать, что в них реализуется первый механизм. Но содержание ДНК в клетках этих штаммов практически не отличается от такового в клетках дикого типа.
Определение содержания GC-nap в ДНК восьми видов бактерий показало, что существует обратная зависимость между GC-содержанием и резистентностью клеток к рентгеновским лучам. В то же время между GC-содержанием и резистентностью к УФ-облучению наблюдается прямая зависимость. Такая корреляция утрачивает какой бы то ни было смысл в случае М. radiodurans, резистентного к обоим типам излучения; однако она может иметь некоторое значение при отсутствии у бактерий эффективных систем репарации. Действительно, ДНК М. radiodurans характеризуется тем же нуклеотидным составом, что и ДНК штаммов Pseudomonas, исключительно чувствительных к ионизирующей радиации (Покровский В.И., 1999).
6.1.1 Механизмы репарации ДНК
В основе радиорезистентности бактерий лежат разнообразные внутриклеточные процессы, участвующие в репарации поврежденной ДНК. Большую ценность для исследования этих процессов представляет наличие хорошо охарактеризованных мутантных штаммов, радиационная чувствительность которых варьирует в чрезвычайно широких пределах.
При помощи генетических скрещиваний были получены двойные и тройные мутанты дрожжей, у которых репаративная активность полностью отсутствует. Сравнительное исследование штамма дикого типа и сверхчувствительных двойных и тройных мутантов S. сеrevisiae показало, что если нормальный штамм довольно легко переносит образование в ДНК почти 16 000 димеров (37% выживания), то двойные и тройные мутанты остаются резистентными в присутствии не более 50 и 1 димера соответственно. Пониженная резистентность таких двойных и тройных мутантов служит убедительным свидетельством в пользу существования различных путей репарации радиационных повреждений.
В зависимости от того, участвует ли видимый свет в модификации повреждений ДНК, репарацию можно подразделить на световую и темновую. Конкретно под световой репарацией понимается феномен фотореактивации, впервые описанный у актиномицетов. Механизм фотореактивации действует только на пиримидиновые димеры. В этом процессе участвует фермент фотореактивации, который связывается с димерами. Образующийся фермент-субстратный комплекс активируется видимым светом, что приводит к мономеризации димеров in situ. Таким образом, летальный эффект УФ-облучения существенно снижается, если облученные клетки подвергаются затем воздействию видимого света с длинами волн от 360 до 420 нм (см. рис. 6.1).
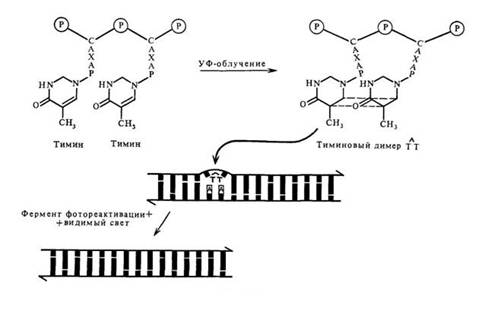
Рис. 6.1 Световая репарация ДНК
Фотореактивация служит мощным инструментом исследования летальных и мутационных повреждений, так как их репарация под влиянием света может быть использована в качестве критерия для решения вопроса о том, обусловлена ли инактивация ДНК образованием пиримидиновых димеров.
К другому типу реактивации клеток видимым светом относится его защитное действие. В этом случае увеличение выживаемости клеток наблюдается при освещении их видимым светом перед УФ-облучением. Этот феномен объясняют тем, что видимый свет индуцирует задержку клеточного деления. В результате такой задержки остается больше времени для репарации повреждений, вызываемых УФ-облучением (см. рис. 6.2).
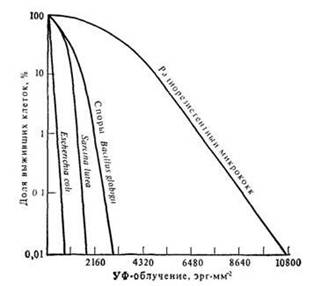
Рис. 6.2 Зависимость выживания клеток бактерий от величины облучения
Под «темновой репарацией» понимают репарацию без участия света. В настоящее время известны две системы такого типа: эксцизионная репарация и пострепликативная рекомбинационная репарация. Репарация первого типа требует присутствия ферментов, которые узнают нарушения структуры ДНК, удаляют затронутые участки, замещая их нормальными нуклеотидными последовательностями, и, наконец, восстанавливают первоначальную структуру ДНК, замыкая полинуклеотидную цепь (см. рис. 6.3).
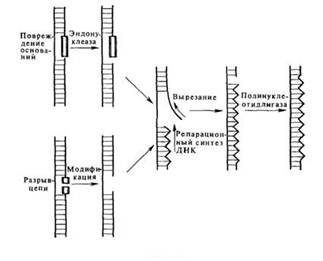
Рис. 6.3Темновая репарация ДНК
Действие разнообразных инактивирующих агентов на клетки может приводить к возникновению в ДНК целого ряда различных повреждений. Детальное изучение системы эксцизионной репарации стало возможным благодаря наличию радиационно-чувствительных мутантов, с помощью которых удалось выделить и охарактеризовать специфические ферменты, принимающие участие в разных стадиях этого процесса. У Е. coli имеется, по крайней мере, четыре таких этапа. На первом этапе происходит разрыв цепи ДНК вблизи повреждения под действием эндонуклеазы, узнающей нарушения структуры ДНК. Такая УФ-специфическая эндонуклеаза была выделена из Micrococcus luteus и Е. coli. За разрывом цепи ДНК следует удаление пиримидиновых димеров, осуществляемое экзонуклеазой. Удаление димеров сопровождается дополнительной деградацией ДНК с образованием брешей, размеры которых варьируют от 20 до 400 нуклеотидов. Затем бреши заполняются с помощью ДНК-полимеразы, использующей в качестве матрицы интактную комплементарную цепь ДНК. Заключительный шаг в этой последовательности событии состоит в восстановлении целостности полинуклеотидной цепи в результате сшивания разрыва лигазой.
Второй тип темновой репарации — пострепликативная рекомбинационная репарация — был впервые описан Говард-Флендерсом. Как указывает само название, эта репаративная система устраняет повреждения в ДНК после того, как произошла ее репликация.
Клеточная система репликации способна «обходить» некоторые из димеров в матричной цепи ДНК, оставляя в растущей цепи бреши, расположенные напротив каждого из них. Число брешей, возникающих таким путем, примерно соответствует числу димеров в ДНК. В результате процесса, сходного с рекомбинацией и включающего обмен между сестринскими нитями, образуется ДНК с двумя интактными цепями. Обнаружено, что при заполнении брешей происходит обмен между облученной родительской цепью ДНК и необлученной дочерней цепью. Установлено, что для образования интактных, не содержащих димеры молекул ДНК, заполнение брешей не обязательно, вместо этого концентрация димеров может просто постепенно снижаться в ходе последовательных циклов репликации ДНК после облучения (Покровский В.И., 1999).
6.1.2 Механизмы резистентности Micrococcus radiodurans
Исключительная резистентность М. radiodurans вызывает особый интерес, исследование кривых выживания выявило очень длинное плечо, выходящее за пределы 1,5 мрад. Этот организм обладает наивысшей резистентностью к гамма-излучению по сравнению со всеми изученными до сих пор микроорганизмами. Для него характерна также необычайная резистентность к УФ-излучению, превышающая резистентность всех исследованных в этом отношении бактерий. Кривая выживания при УФ-облучении состоит из трех компонентов: очень длинного плеча, доходящего до 9000 эрг-мм-2, экспоненциальной части и выраженного «хвоста», который начинается при 25 000 эрг-мм-2 и тянется значительно дальше 50 000 эрг-мм-2. Наличие большого плеча указывает на существование исключительно эффективной клеточной системы репарации.
Имеются различные причины его резистентности к УФ-облучению. В качестве таковых рассматривались экранирование ДНК другими поглощающими соединениями; резистентность, присущая самой ДНК в силу особенностей ее структуры; высокая эффективность репаративных механизмов, а также высокая степень плоидности. Для того чтобы сделать выбор между этими возможностями, исследовались кривые выживания, образование тиминовых димеров как функция дозы УФ-излучения, нуклеотидный состав ДНК и кинетика синтеза ДНК после облучения. В ДНК М. radiodurans при УФ-облучении индуцируется приблизительно втрое меньше тиминовых димеров, чем в ДНК Е. coli. Частично эта разница в димеризации обусловлена различиями в нуклеотидном составе ДНК этих микроорганизмов: у М. radiodurans отношение G + C/A-t-T в 1,6 раза больше, чем у Е. coli (Белозерский, Спирин, 1960). Возможно, имеет значение также разница в поглощении УФ-излучения в расчете на одну клетку. Показано, что эта величина несколько меньше для М. radiodurans, чем для Е. coli. Неизвестно, полностью ли трехкратная разница в димеризации тимина обусловлена различиями в нуклеотидном составе ДНК и поглощении УФ-излучения в расчете на одну клетку. Во всяком случае сам факт уменьшения числа образующихся димеров совершенно недостаточен для объяснения более высокой радиорезистентности М. radiodurans.
При сравнении влияния УФ-облучения на последующий синтез ДНК в клетках М. radiodurans и Е. coli выяснилось, что одинаковая по продолжительности задержка репликации ДНК возникает у них тогда, когда клетки М. radiodurans облучают в дозе, величина которой в 20 раз превышает соответствующую величину для Е. coli. Это различие обусловлено очень высокой эффективностью удаления димеров тимина у М. radiodurans. Димеры тимина вырезаются из клеточной ДНК до возобновления ее репликации. Механизм вырезания пиримидиновых димеров у этой бактерии настолько эффективен, что гибель клеток происходит по каким-то другим причинам, к числу которых может относиться, например, модификация дезоксицитидина и белков.
Повышенная резистентность М. radiodurans к летальному действию облучения явно сопровождается его повышенной резистентностью к индуцированному мутагенезу. Для некоторых радиационно-чувствительных мутантов Е. coll было показано, что характерная для них высокая мутабильность сопутствует их повышенной чувствительности к летальному действию облучения. Ни штамм дикого типа, ни температурно-чувствительный мутант не дают мутаций при УФ-облучении в такой высокой дозе, как 15 000 эрг. Напротив, у Е. coli индуцирование мутаций наблюдается при облучении в дозе 100 эрг.
При облучении в дозе 110 крад, вызывающей значительные физико-химические изменения в ДНК Е. coli В/г, выживание М. radiodurans составляет 100%. Процесс репарации радиационных повреждений у М. radiodurans исключительно точен и не допускает ошибок; этот факт заслуживает особого внимания и может иметь значение для эволюции организмов. Хотя большинство микроорганизмов не способно репарировать двухцепочные разрывы ДНК, М. radiodurans обладает способностью к репарации таких разрывов, индуцируемых гамма-лучами, чем и объясняется его высокая резистентность к ионизирующему излучению. Ионизирующее излучение индуцирует у М. radiodurans освобождение связанной с клеточной поверхностью экзонуклеазы. Это явление можно рассматривать как модель освобождения связанных с мембранами репаративных ферментов после облучения. Более подробное исследование энзимологии репарации ДНК у М. radiodurans было бы полезным для углубления понимания молекулярных основ радиорезистентности (Кашнер Д., 1981).
7. РОЛЬ СТРЕССОСОМ КАК ФАКТОРОВ ВЫЖИВАНИЯ МИКРООРГАНИЗМОВ
О кризисных явлениях в окружающей среде большинству бактерий сигналит особый центр. Этот центр чаще всего является крупной молекулой и назван «стрессосомой». Как правило, бактерия имеет в своём составе около 20 стрессомом, и, хотя ученые знают, что они играют важную роль в генерации клеточного ответа на стрессовые ситуации, сложности в этом процессе не были полностью изучены до сих пор, о кризисных явлениях в окружающей среде большинству бактерий сигналит особый центр. Этот центр чаще всего является крупной молекулой и назван «стрессосомой». Как правило, бактерия имеет в своём составе около 20 стрессомом. Бактерия оказывается в опасной ситуации, например, если температура и соленость среды достигают своего опасного уровня. В таком случае сигнал передается с поверхности клетки внутрь, предупреждая бактерию об угрозе выживанию.
Используя последние достижения электронной микроскопии авторы исследования, результаты которого были опубликованы в журнале Science, отмечают, что стрессосомы, получая сигнал опасности, формируют ответ в виде отделения нескольких белков. Эта белковая структура провоцирует серию сигналов внутри клетки, позволяющих ей адаптироваться и выжить в новой среде.
Стрессосомы клетки являются важнейшими элементами для защиты клетки, поскольку они позволяют очень быстро реагировать на опасность. Цепные реакции, происходящие в результате их активации, позволяют бактерии адаптироваться к изменениям в её окружающей среде почти мгновенно. Каскад событий внутри бактериальной клетки, который возникает вследствие сигнала стрессосомы, заставляет конкретные гены внутри клетки усиливать процесс трансляции. Это означает, что некоторые гены внутри клетки включаются в момент опасности и вызывают увеличение количества определённых белков. Именно такие изменения в клетке позволяют ей выживать во враждебных ей условиях (Марлз Дж., Грант Т., 2008).
ЗАКЛЮЧЕНИЕ:
Таким образом, в завершение данного курсового проекта, можно подвести обобщающие выводы.
Были рассмотрены причины, по которым одни микроорганизмы способны размножаться при пониженных температурах, а другие даже нуждаются в таких условиях. Модификация мембранных липидов, а следовательно, и изменения в функционировании мембран представляют собой важный аспект температурной адаптации. Этот аспект особенно интересен в свете рассмотрения форм жизни, которые могли бы существовать на гигантских планетах, где условия, более или менее приемлемые для жизни, возможны лишь в газообразных областях.
Способность некоторых микроорганизмов жить при высоких температурах уже давно привлекла внимание биологов. Температурная адаптация микроорганизмов обусловлена изменениями в скоростях метаболизма, а также в структуре мембран, рибосом и отдельных белков. Наиболее важными для адаптации к высоким температурам являются изменения в структуре белков. При тех высоких температурах, при которых растут термофилы, многие их ферменты сохраняют как активность, так и регуляторные свойства.
Многие микроорганизмы способны размножаться в интервале значений рН, в котором их внутриклеточные ферменты не функционируют. Микроорганизмы могут существовать при концентрациях водородных ионов, различающихся на несколько порядков; отдельные микроорганизмы растут при рН 10 и даже при более высоких. Несмотря на то что рН окружающей среды может меняться, внутри своих клеток эти организмы поддерживают постоянную кислотность. Структуры на поверхности клеток у таких организмов должны быть приспособлены к крайним значениям рН.
Экстремальные галофилы занимают особое место среди микроорганизмов, существующих в экстремальных условиях, поскольку они представляют собой пример полной (и внешней, и внутренней) адаптации к очень высоким концентрациям солей, а также потому, что они обладают уникальными биохимическими свойствами. С недавних пор стало ясно, что организмы, живущие при высоких концентрациях растворенных веществ или способные размножаться в широком диапазоне концентраций, представляют собой крайне увлекательный объект исследования.
Многие микроорганизмы сохраняют жизнеспособность в течение долгого времени в отсутствие воды и начинают размножаться, как только она снова становится доступна для них. Хотя для своего размножения микроорганизмы нуждаются в определенном уровне содержания воды, она не требуется им для выживания.
Токсичность тяжелых металлов представляет собой проблему скорее для человека, чем для микроорганизмов, которые научились по-разному приспосабливаться к таким веществам. Микроорганизмы способны осуществлять трансформацию тяжелых металлов в окружающей среде: выщелачивать металлы из руд в кислых рудничных стоках, изменять валентность металлов, как, например, при трансформации ртути в более или менее токсичные формы, а также при образовании таких особых форм скоплений металлов, как марганцевые конкреции.
Микроорганизмы сильно отличаются друг от друга по своей устойчивости к радиации. Многие из них способны выдерживать дозы радиации, летальные для других форм жизни. Подобная устойчивость вызвана рядом факторов, наиболее важным из которых представляется способность микроорганизмов к репарации их ДНК, поврежденных облучением. Многообразие способов, при помощи которых микроорганизмы противостоят радиации, может сделать их последними обитателями на Земле или, напротив, первыми поселенцами на Земле, разрушенной атомной войной.
СПИСОК ЛИТЕРАТУРЫ
1. Асонов Н.Р. Микробиология / Н.Р. Асонов. – М.: Колос, 1980. – 312 с.
2. Камшилов М. М. Эволюция биосферы / М.М. Камшилов. - М.: Наука, 1974. - 254 с.
3. Кашнер Д. Жизнь микробов в экстремальных условиях. Пер. с англ. / Д. Кашнер. – М.: Мир, 1981. – 511 с.
4. Логинова Л.Г. Новые формы термофильных бактерий / Л.Г. Логинова, Л.А. Егорова. - М.: Наука, 1977. - 175 с.
5. Лях С.П. Адаптация микроорганизмов к низким температурам / С.П. Лях. - М.: Наука, 1976. – 160 с.
6. Марлз Дж., Грант Т., Делюмье О. Молекулярное строение стрессосом // Science, 2008. №3 C. 92 – 96
7. Радчук Н.А. Ветеринарная микробиология и иммунология / Н.А. Радчук, Г.В. Дунаев, Н.М.Колычев и др. – М.: Агропромиздат, 1991. – 383 с.
8. Покровский В.И. Микробиология / В.И. Покровский, О.Н. Поздеев. – М.: ГЭОТАР, 1999. 1200с.
9. Бухарин О.В. Механизмы выживания бактерий / Бухарин О.В., Гинцбург А.Л., Романова Ю.М. и др. - М.: Медицина, 2005. – 367 с.
10. Заварзин Г.А. Природоведческая микробиология / Заварзин Г.А., Колотилова Н.Н. – М.: Книжный дом «Университет», 2001. – 256 с.
© 2010 Интернет База Рефератов