
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Курсовая работа: Папоротники Пензенской области
Курсовая работа: Папоротники Пензенской области
Пензенский Государственный Педагогический
Университет им. В.Г. Белинского
Курсовая работа
на тему:
Папоротники Пензенской области
Студента группы Бх-21
Ащина Никиты
Пенза-2006
Введение
Папоротниковидные относятся к числу наиболее древних групп высших растений. По своей древности они уступают только риниофитам и плауновидным и имеют приблизительно один геологический возраст с хвощевидными. Но в то время как риниофиты давно вымерли, а плауновидные и хвощевидные играют в современном растительном покрове Земли очень скромную роль их число их видов невелико, папоротники продолжают процветать. Хотя сейчас они играют несколько меньшую роль, чем в прошлые геологические периоды, тем не менее и в настоящее время насчитывается около 300 родов и более 10 000 видов папоротников.
Папоротники распространены очень широко, фактически по всему земному шару, и встречаются в самых различных местообитаниях, начиная с пустынь и кончая болотами, озерами, рисовыми полями и солоноватыми водами. Но наибольшее их разнообразие наблюдается во влажных тропических лесах, где они обильно произрастают не только на почве под деревьями, но и в качестве эпифитов на стволах и ветвях деревьев, часто в очень большом количестве. В результате приспособления к столь различным условиям среды у папоротников выработались очень разные жизненные формы и возникло очень большое разнообразие во внешней форме, внутреннем строении, физиологических особенностях и размерах. По своим размерам папоротники варьируют от тропических древовидных форм до крошечных растеньиц длиной всего лишь в несколько миллиметров.
На территории Пензенской области произрастают несколько видов Папоротниковидных. И целью данной курсовой работы является исследование всех видов папоротников, которые произрастают на территории Пензенской области.
Глава 1. Общая характеристика отдела ПАПОРОТНИКООБРАЗНЫЕ (РТЕRIDOPHYTA, ИЛИ POLYPODIOPHYTA)
1.1 Особенности строения и размножения видов отдела Папоротникообразные
Подобно плауновидным и членистым папоротникообразные являются древнейшей группой, известной с девонского периода. Расцвета они достигли в конце палеозойской-начале мезозойской эры, когда были представлены большим разнообразием жизненных форм и распространены на всех континентах земного шара. Наибольшую роль в сложении растительного покрова Земли играли крупные древовидные папоротники, входившие в состав каменноугольных лесов. В настоящее время папоротники насчитывают более 10 тыс. видов и 300 родов.
Для папоротникообразных характерно сочетание ряда признаков, из которых главнейшими являются макрофилия, отсутствие камбия и отсутствие стробилов. Под макрофилией понимаются прежде всего относительно крупные размеры листьев, которые часто называются вайями. В отличие от плауновых и членистых листья папоротников имеют более сложное морфологическое и анатомическое строение; они состоят из основания - филлоподия, черешка и листовой пластинки, часто многократно рассеченной, с густой сетью жилок. Для листовой пластинки наиболее характерен длительный рост ее вершины. Каждому листу в стебле соответствует листовая лакуна (листовой прорыв). Этот комплекс признаков отражает синтеломное происхождение листьев папоротников, т.е. возникновение их из системы вегетативных, спороносных либо смешанных теломов, что подтверждается палеонтологическими данными.
Ныне живущие папоротники распространены на всех континентах, встречаясь в самых разнообразных экологических условиях. Однако ведущая роль и по количеству видов, и по разнообразию жизненных форм принадлежит папоротникам влажных тропических и субтропических лесов, где многие семейства находятся в расцвете формообразования. Условия обитания наложили существенный отпечаток на их анатомо-морфологические и биологические особенности. Современные папоротники представлены в основном травянистыми растениями, а древовидные составляют незначительное количество. Папоротники умеренных областей, за исключением сальвинии плавающей (Salvinia natans), - многолетние травянистые растения с подземными длинными или короткими корневищами. У длиннокорневищных папоротников - у орляка (Pteridium aqualinum), голокучника Линнея (Gymnocarpium dryopteris) и др. - длина междоузлий измеряется сантиметрами, поэтому над землей листья располагаются на большом расстоянии друг от друга. У большинства же папоротников на коротких корневищах формируется розетка листьев, как правило, отмирающих осенью, при этом их расширенные основания длительное время остаются на корневище, образуя плотный толстый чехол вокруг довольно тонкого, диаметром до I см, стебля.
В зоне влажных тропических и субтропических лесов разнообразие жизненных форм чрезвычайно велико. В тенистых лесах много наземных видов, большинство из них имеют длинные стелющиеся побеги, реже встречаются виды с короткими прямостоячими побегами. Самые мелкие наземные папоротники из рода трихоманес (Тrichomanes) имеют длину от 3-4 мм до 2-4 см, а самые крупные виды ангиоптериса(Angiopteris), образующие часто густые заросли, имеют клубневидные стебли диаметром до 1 м. На них располагаются листья длиной 5-6 м с длинными крепкими черешками и сильно рассеченными листовыми пластинками. Однако наиболее многочисленны и разнообразны эпифитные папоротники, особенно в тропических лесах Старого Света. Вездесущие эпифиты встречаются на моховых подушках, на стволах и в кронах деревьев. Многие приземные эпифиты, в основном из семейства гименофилловых (Hymenophyllaceae), находятся в условиях избыточного увлажнения; их нижние прозрачные листья толщиной в 1-3 слоя клеток лишены устьиц и поглощают атмосферную влагу всей поверхностью. Эпифиты, живущие в кронах и на стволах деревьев, находятся в условиях дефицита влаги, в связи с чем имеют листья плотные, кожистые либо сильноопушенные. Один из замечательных папоротников-эпифитов, придающий своеобразный облик лесам Старого Света, - асплениум гнездовый, или птичье гнездо (Asplenium nidus), издали создает впечатление огромных птичьих гнезд (рис. 1). Его короткие толстые побеги прочно закрепляются на стволах и ветвях деревьев с помощью многочисленных переплетающихся и сильно опушенных корней. На вершинах побегов формируются изумительной красоты розетки кожистых листьев, достигающих иногда длины до 2 м.

Рис. 1 - Жизненные формы папоротников: А, Б - эпифитные папоротники - асплениум гнездовый (Asplenium nidus); платацериум (Р1аcerium); В - листовая лиана лигодиум (Lygodium); Г - древовидная форма
Вся масса листьев и корней способна накапливать гумус и поглощать влагу из атмосферы, обеспечивая себе и питание, и водоснабжение. Во всех тропических лесах широко распространены виды рода платицериума, или оленьего рога (Р1аtусеrium). Их короткие стебли также прикрепляются к коре деревьев многочисленными корнями. Вегетативные листья имеют вид плоских округлых пластинок, которые своими основаниями плотно прижаты к стволу, а их вершины отстоят от стебля, образуя нишу в виде кармана. В ней скапливаются опадающие и перегнивающие кора деревьев и листья самого растения, т.е. они сами формируют себе почву. У платицериума большого (Р. grande) в такой нише, глубиной более 1 м, может скапливаться до 100 кг почвы; под их тяжестью деревья-носители иногда выворачиваются с корнями. Несколько позднее возникают вегетативные или спороносные листья, дихотомически или пальчато-ветвящиеся, за что растения и получили название олений рог.
Аналогичные ниши в виде карманов могут образовывать плоские стебли некоторых папоротников, например малайский вид многоножки (Ро1уроdium imbricatum). Однако наряду с высокоспециализированными описанными формами существует немало мелких эпифитов и эпифиллов, относящихся к родам схизея (Schizаеа) и трихоманес (Тrichomanes). У одних видов мелкие листья покрыты гигроскопическими волосками, снижающими испарение, а во влажные периоды поглощающие влагу из атмосферы. Другие виды, лишенные волосков, способны в наиболее сухое время суток переходить в состояние анабиоза. Значительно меньше папоротников лиан, причем они менее специализированны, чем эпифиты. Одни просто опираются на стволы деревьев своими тонкими длинными стеблями, у других имеются либо изогнутые черешки, либо шипы на листьях, либо придаточные корни на стеблях. Наибольший интерес представляет род лигодиум (Lygodium), имеющий уникальную жизненную форму листовой лианы. Вдоль длинного стелющегося побега с нижней стороны отходят корни, а на верхней стороне двумя рядами располагаются очень своеобразные многократно перисторассеченные листья; их черешки способны к весьма длительному нарастанию, достигая иногда длины 30 м. Обвиваясь вокруг стволов деревьев, они выносят вверх доли листа ближе к свету.
Весьма своеобразны древовидные папоротники, к которым относятся 8 родов. Многие из них, например диксония (Dicksonia), циатея (Суаthеа), приурочены к горным районам тропической и субтропической зон, где они образуют изящные рощи.
Большинство древовидных папоротников достигает высоты от 5-6 до 10 м, редкие экземпляры имеют максимальные размеры 20-25 м при диаметре ствола 50 см. Стволы их, как правило, не ветвятся и образуют на вершине раскидистую ажурную крону из перистых листьев длиной 2-3 м (у циатеи до 5-6 м). У многих видов в кроне выявляются 3 категории листьев - молодые, с еще не развернувшимися улитками, листья направлены вверх; срединные расположены горизонтально, а увядающие листья, отгибаясь вниз, образуют своеобразную «юбочку». При опадании листьев часто на стебле "остаются их основания и нижние части сильно склерофицированных черешков, которые одевают стебель в виде футляра. Кроме того, под каждым листом формируются корни, одни из которых достигают поверхности земли, а другие остаются воздушными. Стеблевые корни также пронизаны склеренхимными пучками, что придает им большую прочность. Черешки, направленные вверх, и корни, направленные вниз, переплетаясь между собой, образуют вокруг стебля прочный цилиндр наподобие панцирной сетки, выполняющий опорную функцию. При значительной высоте растений это оказывается важным обстоятельством, так как древовидные папоротники представляют собой, по существу, гигантские травы.
Поскольку у папоротников камбий отсутствует, у них нет и вторичной древесины; механическая прочность достигается за счет склеренхимной обкладки вокруг проводящих пучков; лишь иногда внешняя кора состоит из механической ткани. Поэтому внешний листо-корневой цилиндр выполняет основную опорную функцию. По мере старения растения основание его ствола отмирает и разрушается, однако ствол не падает, так как он, как на ходулях, удерживается свисающими корнями. Провести четкую границу между травами и древовидными формами невозможно. В пределах одного вида размеры могут варьировать от нескольких дециметров до нескольких метров, что в значительной степени определяется почвенными и температурными условиями. Даже такой краткий перечень жизненных форм говорит о большом морфологическом разнообразии папоротников; это касается листьев, стеблей и побегов в целом. Морфологическая характеристика папоротников часто сталкивается с терминологическими трудностями, так как термины и понятия, разработанные для цветковых, не всегда приемлемы для папоротников. Это прежде всего относится к понятию корневища папоротников, которое в ходе онтогенеза может менять свою природу. У изученных видов корневищных папоротников зигота при делении образует 4 клетки; из одной возникает гаустория, из второй - корень, из третьей - лист, из четвертой - стебель, т.е. стебель, корень и лист - это равноценные гомологичные органы этих папоротников. Чаще всего зародышевый лист и корень опережают в своем развитии стебель, поэтому происходит формирование листа с корнем у основания. В основании черешка первого листа закладывается меристематический бугорок, формирующий новый лист с корнем у основания. Следующий лист возникает из бугорка, расположенного в основании черешка предыдущего листа. Основания всех вновь возникающих листьев и корней в совокупности формируют корневище, уникальное для ныне живущих высших растений. Его иногда называют филлогенным, т.е. развивающимся из листьев. Однако в ходе онтогенеза филлогенное корневище сменяется обычным; при этом мер истом этически и бугорок - точка роста начинает формировать стебель с зачатками листьев. У длиннокорневишных папоротников на вершине, в непосредственной близости от верхушечной (апикальной) инициальной клетки, вычленяется бугорок из меристематических клеток. Из него может развиться либо боковое корневище, либо лист, что также указывает на гомологичность этих органов. Вершина корневища длиннокорневищных папоротников может быть покрыта многочисленными чешуйками или оставаться совсем голой. У короткокорневищиых форм на самой вершине закладываются особые листья с недоразвитой листовой пластинкой и хорошо развитым основанием - филлоподием, они обеспечивают дополнительную защиту зимующих почек. Ветвление растения может осуществляться не только за счет стеблевых почек, но и почками, возникающими на черешках листьев или на листовых пластинках. Часто такие листовые почки сразу формируют молодые розетки, которые опадают и осуществляют вегетативное размножение; такие растения называют живородящими. У некоторых папоротников, например у нефролеписа клубненосного (Nephrolepis tuberosa) из стеблевых почек развиваются тонкие столонообразные корневища, лишенные листьев и покрытые кожистыми чешуйками. Выходя на поверхность земли, они формируют новую розетку. Кроме того, на столонах возникают клубневидные боковые ответвления, осуществляющие вегетативное размножение.
Весьма специфичны листья папоротников - вайи. Для них наиболее характерен длительный рост вершины, проявляющийся в образовании улитки (исключение составляют ужовниковые), и густая сеть обильно ветвящихся жилок. Развитие листьев под землей часто длится несколько лет, а над землей завершается в течение 1 - 1,5 недель. У некоторых видов, например у адиантума (Аdianthum), камптосоруса (Сamptosorus), листья проявляют стеблевую природу - их рахисы вытягиваются в стеблевидную плеть и, достигнув поверхности земли, укореняются с образованием новой розетки. У описанного выше лигодиума рахис листа своим поведением также очень напоминает стебель. Основания листьев, черешки и листовые пластинки у многих видов покрыты чешуйками, которые рассматриваются иногда как микрофиллы, т. е. листья энационного происхождения. Разнообразие их форм, размеров, цвета составляет важный систематический признак. Еще более разнообразны листовые пластинки. Наиболее типичны листья дважды, трижды- и более перисторассеченные, реже встречаются листья пальчаторассеченные и еще реже - дихотомически рассеченные. Центральную часть листовой пластинки, представляющую продолжение черешка, называют рахисом, а боковые доли первого и последующих порядков - соответственно перьями и перышками. Наряду с рассеченными листьями в разных семействах имеются цельные листья, сформировавшиеся в результате полного слияния боковых долей. Характерно, что во влажных тропических лесах цельными листьями обладают, как правило, мелкие папоротники, а все крупные папоротники имеют рассеченные листовые пластинки. Это, по-видимому, стоит в связи с характером тропических ливней, при которых мощные потоки воды свободно проходят сквозь рассеченную листовую пластинку, не нарушая ее целостности. Не менее разнообразно жилкование листьев от наиболее примитивного открытого до более совершенного - сетчатого.
У большинства видов папоротников листья совмещают 2 функции - фотосинтеза и спороношения (рис. 2), однако у многих видов наблюдается диморфизм листьев - одни выполняют функцию фотосинтеза, а другие - только спороношения, например у страусника, трихоманеса. У некоторых видов папоротников (у ужовниковых, у осмунды) имеется диморфизм частей листа, при котором одна часть выполняет функцию фотосинтеза, а другая часть - спороношения. Палеонтологический материал показывает, что все 3 типа листьев существовали уже в раннем палеозое и формировались независимо друг от друга.
Ход развития корней папоротников до сего времени мало изучен. Как уже отмечалось, при формировании зародыша зародышевый корень закладывается одновременно со стеблем и листом, при этом смещается гаусторией вбок. В дальнейшем у одних видов он развивается одновременно с листом, а у других видов несколько задерживается в росте и появляется позднее, поэтому производит впечатление придаточного корня.
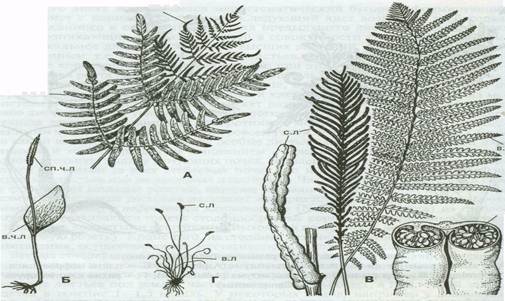
Рис. 2 - Спороносные листья папоротников: А - осмунды (Оsmunda); Б - ужовника(Оphioglossum); В - страусника (Маtteuccia); Г - трихоманеса (Тrichomanes); Д - схизеи (Schizaeae); в. ч. л - вегетативная часть листа; сп. ч. л - спороносная часть листа; в. л - вегетативный лист; с.л - спороносный лист
У одних видов корни могут появиться на поверхности побега в год его заложения, а у других видов - лишь через 1-2 года. От настоящих придаточных корней семенных растений корни папоротников (так же, как у плаунов и хвощей) отличаются тем, что не могут закладываться на уже сформированных частях побегов. Корни папоротников живут 3-4 года. У некоторых видов, например у нефролеписа, корни, загибаясь вверх, способны превращаться в олиственные побеги.
Анализ вегетативных органов папоротников выявляет способность превращения одного органа в другой, т.е. указывает на то, что дифференциация на органы у них не всегда жестко закреплена генетически. Это находится в прямой связи с древностью папоротников. Сходство анатомического строения стеблей, черешков листьев и корней также указывает на единство происхождения всех вегетативных органов.
У большинства современных видов папоротников стебли имеют диктиостелу, однако ход морфогенеза у разных видов различен. У многих изученных папоротников стебель молодого растения построен по типу протостелы, затем формируется эктофлоическая или амфифлоическая сифоностела, а с началом образования листовых лакун происходит формирование диктиостелы. Она представляет собой цилиндр, в центре которого расположена паренхима сердцевины, затем следуют внутренняя флоэма, ксилема и внешняя флоэма, пронизанные паренхимой листовых лакун. На поперечном срезе листовые лакуны представлены сердцевинными лучами, а между ними по кольцу располагаются концентрические проводящие пучки с ксилемой в центре и флоэмой по периферии. При мацерации все живые ткани сердцевины, сердцевинных лучей и флоэмы разрушаются и остается лишь ксилема в виде сетчатого цилиндра, отсюда и название диктиостела - сетчатая стела. Флоэма одночленная, состоит только из ситовидных клеток. Ксилема сложена в основном лестничными, частично кольчатыми трахеидами. Наряду с диктиостелой у некоторых древних видов на протяжении всей жизни сохраняется протостела (у лигодиума, глейхении), эктофлоическая сифоностела (у циатеи) или амфифлоическая сифоностела (у видов глейхении, у диптериса). Все это позволяет сделать вывод о направленности эволюции стелы от протостелы через сифоностелу к диктиостеле, что подтверждается палеоботаническим материалом. У пермских папоротников описана протостелическая структура, у триасовых - в основном сифоностелическая, у большинства современных - диктиостела, при которой осуществляется наибольший контакт проводящих тканей с живой паренхимой, а следовательно, улучшается водоснабжение. Размножение папоротников осуществляется в основном за счет спор. Большинство видов папоротников - равноспоровые растения; количество разноспоровых видов невелико. Для всех папоротников характерно отсутствие специализированных спороносных побегов - стробилов. У большинства папоротников спорангии сгруппированы в сорусы; у мараттиевых, срастаясь между собой, они образуют синангии (рис. 3). У наиболее примитивных видов одиночные спорангии располагаются по краям листьев либо на вершинах их лопастей, при этом каждый спорангий снабжается самостоятельной жилкой. Это весьма напоминает верхушечное расположение спорангиев на концах васкуляризованных теломов у риниофитов.

Рис. 3 - Типы размещения спорангиев и сорусов: А - расположение одиночных спорангиев: 1 - верхушечное по отношению к долям листа у даваллиевых (Davalliacеае); 2 - краевое у трихоманеса (Тrichomanes); Б - расположение сорусов: 1 - краевое у гименофилла (Нуmophillum); 2 - на нижней стороне листа у кочедыжника ломкого (Суstopteris fragilis); В - синангии у мараттиевых (Магаttiacеае): 1 - Аngiopteris; 2 - Магattiа; 3 - Сhristensiа; Г -перемещение спорангиев на нижнюю поверхность листа в ходе морфогенеза у схизеи (Schizaea); сп -спорангий
У большинства же папоротников спорангии или сорусы располагаются на нижней поверхности листьев. Чаще всего они приурочены к главной и боковым жилкам, реже (например, у орляка) - близ края листа. Расположение спорангиев на нижней стороне листа оказывается биологически выгодным: во-первых, обеспечивается надежная защита спорангиев во время их созревания и одновременно не снижается интенсивность фотосинтеза. Во-вторых, обеспечивается более равномерное рассеивание спор, в треть их создается возможность большего продуцирования спорангиев на всей поверхности листа, чем только по краю. Ход морфогенеза спороносных листьев у некоторых видов рода схизеи показывает возможный путь перемещения спорагниев на нижнюю поверхность листа в ходе филогенеза. На ранних этапах развития листа схизеи спорангии закладываются по его краям, но в результате более быстрого развития тканей верхней стороны листа происходит загибание его краёв вниз. В результате оказываются на нижней стороне листа. Место прикрепления спорангии к листу называется плацентой. В ходе эволюции происходило разрастание плаценты, и она приобретала продолговатую или шаровидную форму, что увеличивало площадь возможного размещения спорангиев. Кроме того, на плаценте спорангии располагаются в определенной последовательности, a следовательно, и созревание и вскрывание их происходит неодновременно. Растянутость созревания спор во времени гарантирует более надежное их распространение. Увеличение количества спорангиев на плаценте достигается также увеличением длины ножки спорангия. Разрастание плаценты и увеличение длины ножки коррелятивно не связаны, эти два процесса происходили в разных систематических группах независимо друг от друга.
Большое биологическое значение имеет защита сорусов в период их созревания. В простейшем случае сорусы прикрываются загибающимся вниз краем листа. У более специализированных видов формируется особое покрывало индузиум, возникающее в результате локального разрастания плаценты либо поверхностных тканей листа. По характеру заложения и строению стенки спорангия папоротники подразделяются на лепто- и эуспорангиатн ы е. У эуспорангиатных папоротников спорангий возникает из группы клеток и имеет многослойную стенку, у лептоспорангиатных он возникает из одной клетки и имеет однослойную стенку. И тот и другой тип спорангиев обнаружены у древних папоротников. Механизмы вскрывания спорангиев также чрезвычайно многообразны. В простейшем случае на вершине спорангия имеется небольшой участок сильно утолщенных клеток - пора. При подсыхании спорангия происходит его разрыв на контакте тонкостенных и толстостенных клеток. Однако уже у древних папоротников возникло так называемое кольцо - полоска из толстостенных клеток. У представителей одних семейств оно занимает горизонтальное положение, у других - косое, у третьих - вертикальное. Примитивным считается кольцо сплошное, или замкнутое; более совершенным - неполное, при котором часть клеток кольца остается неутолщенным (так называемое устье); по нему происходит разрыв стенки. Примером может служить спорангий щитовника мужского (Driopteris filix-mas), имеющий форму двояковыпуклой линзы и расположенный на длинной ножке. Однослойная стенка спорангия состоит из крупных тонкостенных клеток. По гребню спорангия, начиная от ножки, проходит кольцо, состоящее на 2/3 из толстостенных клеток, а 1/3 приходится на устье. Клетки кольца имеют утолщения на трех стенках - на двух радиальных и на внутренней - тангентальной. При подсыхании клетки кольца теряют воду, что приводит к уменьшению их объема. Большая сила сцепления втягивает тонкую внешнюю тангентальную стенку внутрь клетки, и она прогибается, притягивая при этом радиальные стенки друг к другу. Это приводит к уменьшению длины окружности кольца и создает большое натяжение в области устья. Происходит
стремительный разрыв стенки спорангия, а кольцо при этом выворачивается наружу, разбрасывая споры. По мере дальнейшего испарения воды тонкая тангентальная стенка касается внутренней, сила сцепления исчезает, и кольцо пружинисто возвращается в свое прежнее положение, разбрасывая остатки спор.
У представителей примитивных семейств спорангии крупные, немногочисленные, содержащие большое количество спор (8-15 тыс.). В продвинутых семействах многочисленные последовательно возникающие спорангии мелкие, содержат, как правило, от 64 до 16 спор. Это обеспечивает автономность спорангиев и увеличивает надежность сохранения спор.
Споры могут находиться в покоящемся состоянии от нескольких недель до нескольких лет и даже десятков лет. Для их прорастания необходимы влага, положительные температуры, определенная кислотность почв, интенсивность и качество света, специфичное для каждого вида.
Существуют разные варианты прорастания спор и хода морфогенеза гаметофитов, которые оказываются характерными не только для отдельных семейств и даже родов. Поэтому некоторые птеридологи при классификации папоротников базируются на особенностях развития и строения гаметофитов.
Гаметофиты большинства равноспоровых папоротников ведут наземный образ жизни, осуществляя автотрофное питание. Продолжительность их жизни, как правило, исчисляется несколькими месяцами, и лишь у отдельных наиболее примитивных видов гаметофиты живут несколько лет (иногда до 10-15). При общей тенденции к сокращению продолжительности жизни гаметофиты некоторых видов папоротников стали вторично многолетними. Формирование гаметофита в значительной степени зависит от спектрального состава света - в синих лучах происходит длительное нарастание нити, а в красных лучах формируются пластинчатые формы. Вначале рост однослойной пластинки осуществляется за счет краевых клеток, а затем на вершине таллома обособляется одна инициальная клетка, которая формирует верхушечную меристему. При этом пластинка (размером в несколько миллиметров) приобретает сердцевидную форму с подушковидной центральной частью. Физиологически активная меристема выделяет специфический гормон, близкий к ростовым гормонам, - антеридиоген; он стимулирует формирование антеридиев на близрасположенных заростках. На нижней стороне пластинки у ее основания среди многочисленных ризоидов развиваются антеридии. Немного позднее у вершины пластинки на многослойной подушке формируются архегонии. Неодновременность развития антеридиев и архегониев способствует перекрестному оплодотворению. Как показали опыты, при большом скоплении заростков концентрация антеридиогена в окружающей среде настолько возрастает, что на вновь формирующихся молодых гаметофитах антеридии могут возникать очень рано, иногда на стадии 2-3-клеточной нити. Высокая концентрация антеридиогена приостанавливает дальнейшее развитие гаметофита, а следовательно, и формирование архегониев, поэтому он остается однополым мужским. В отсутствие антеридиогена при благоприятных условиях происходит быстрое развитие заростка, завершающееся фазой сердцевидной пластинки с хорошо развитой меристемой. На заростке, минуя стадию образования антеридиев, формируются архегонии, т.е. он оказывается однополым женским. При слабом воздействии антеридиогена гаметофит проходит все стадии морфогенеза, формируя сначала антеридии, а затем архегонии, - возникает обоеполый заросток. В природе взаимные влияния гаметофитов друг на друга оказываются сложнее и труднее поддаются наблюдениям.
Описанный тип морфогенеза гаметофита хотя и наиболее распространенный, но не единственный. У схизейных и некоторых гименофилловых заростки в течение всей жизни сохраняют обильно ветвящуюся нитевидную форму, поэтому антеридии и архегонии возникают у них без видимой закономерности на боковых ветвях. У мараттиевых зеленые заростки размером 2-3 см многослойные, мясистые, живут по нескольку лет. У ужовниковых, некоторых схизейных и гдейхениевых гаметофиты клубневидной или червеобразной формы ведут подземный образ жизни, питаясь микотрофно, поэтому они бесцветные. Однако попадая на поверхность земли, они зеленеют, из чего можно заключить, что их подземное существование вторично. Антеридии примитивных семейств крупные, с множеством сперматозоидов, а у более продвинутых представителей они мелкие, содержащие до 32 сперматозоидов. Архегонии у всех папоротников однотипные, погруженные в ткань гаметофита. Ход прорастания зиготы и формирование молодого растения описаны выше.
От типичного цикла развития папоротников имеются отклонения. Иногда спорофит может развиваться апогамно, т.е. без оплодотворения, из одной или группы вегег тативных клеток гаплоидного гаметофита. В этом случае спорофит оказывается гапло-идным, а споры формируются без редукционного деления. В других случаях гаметофит возникает апоспорически не из гаплоидной споры, а из дишюидных тканей спорофита (из клеток спорангия, листа и т.д.), т.е. он оказывается диплоидным. В этом случае диплоидная яйцеклетка без оплодотворения развивается в новый спорофит.
Вегетативное размножение широко распространено среди папоротников. Часто оно осуществляется с помощью выводковых почек, возникающих на листьях, стеблях и корнях. В последнем случае корни, загибаясь, выходят на поверхность земли и на вершине формируют почку. Нередко на листьях и стеблях развиваются длинные плети, на которых возникают почки.
Классификация всего отдела папоротниковидных и особенно вымерших форм испытывает существенные затруднения. В ее основу могут быть положены разные признаки - анатомическое и морфологическое строение вегетативных органов спорофита, особенности спороношения (строение спорангиев и их размещение), характер формирования и строение гаметофитов. Наибольшей ревизии подверглись древние вымерщие группы папоротников - первопапоротники. Такие таксоны, как анейрофитовые (Аneurophytopsida) и археоптерисовые (Archaepteridopsida), по наличию ярко выраженной вторичной ксилемы и наличию окаймленных пор на стенках точечных трахеид отнесены к праголосеменным растениям (Ргоgymnospermae). В основу классификации ныне живущих папоротников положено строение стенки спорангия в сочетании с рядом морфологических признаков.
Классификация древних папоротников еще более осложняется тем, что в палеозое имелись синтетические виды, которые объединяли в себе признаки различных таксонов. Перекомбинация признаков, которая вообще была характерна для большинства раннепалеозойских растений, делает выделение таксонов условным.
1.2 Классификация папоротников Пензенской области
I. Подкласс OPHIOGLOSSIDAE - ОФИОГЛОССИДЫ
Семейство Ophioglossaceae – УЖОВНИКОВЫЕ(2/3)
Род Ophioglossum L. – Ужовник
O. vulgatum L. – У. Обыкновенный
Род Botrychium Sw. – Гроздовник
B. lunaria(L). Sw – Г. Полулунный
B. multifidum (S. G. Gmel.) Rupr. – Г. Многораздельный
II. Подкласс POLYPODIDAE – ПОЛИПОДИДЫ
Семейство Onocleaceae – ОНОКЛЕЕВЫЕ(1/1)
Род Matteuccia Tod. – Страусник.
M. struthiopteris (L) Tod. – С.обыкновенный
Семейство Aspleniaceae - Костенцовые (-/-)
Семейство Athyriaceae – Кочедыжниковые(4/4)
Род Athyrium Roth – Кочедыжник.
A.filix-femina (L.) Roth – К.женский.
Род Diplazium Sw. – Диплазий.
D.sibiricum (Turcz.ex G. Kuntze.) Kurata. – Д.сибирский.
Род Cystopteris Bernh. – Пузырник, или Цистоптерис.
С. Fragilis (L.) Bernh. – Ц. ломкий.
Род Gymnocarpium Newm. – Голокучник.
G.dryopteris (L.) Newm. – Г.обыкновенный.
Семейство Aspidiaceae – Щитолистниковые(1/3)
Род Driopteris Adans. – Щитолистник.
D.carthusiana (Vill.) H.P. Fuchs. – Щ. Картузиса или игольчатый.
D.cristata (L.) A. Gray – Щ. гребенчатый.
D. filix-mas (L.) Schott – Щ. мужской.
Семейство Thelypteridaceae – Телиртерисовые (2/2)
Род Thelyrteris Schmidel Телиптерис.
Th.palustris Schott - Т.болотный.
Род Phegopteris Fee – Фегоптерис.
Ph. Connectilis (Michx.) Watt - Ф.связывающий.
Семейство Hypolepidaceae – Гиполеписовые(1/1)
Род Pteridium Scop. – Орляк.
P.aquilinum (L.) Kuhn ex Decken - О.обыкновенный.
Семейство Polypodiaceae – Многоножковые(-/-)
Семейство Marsileaceae – Марсилиевые(-/-)
Семейство Salviniaceae – Cальвиниевые(1/1)
Род Salvinia Seguier – Cальвиния.
S.natans (L.) All. – C.плавающая.
1.3 Ключ для определения родов и видов отдела Папоротникообразные
Насчитывает 3 рода и 10 видов, распространённых в Северно м полушарии. В Средней России только один род Matteuccia Todaro -- Страусник с одним видом 1. М. struthiopteris (L) Todaro (Struthiopteris filicastrum All.) - С. обыкновенный
СЕМЕЙСТВО ATHYRIACEAE - КОЧЕДЫЖНИКОВЫЕ
Насчитывает 25 родов и около 600 видов, распространённых по всему земному шару. В Средней России 3 рода.
Сорусы от продолговатых до линейных; покрывал ьце прикреплено вдоль сорусов своим широким краем. Сорусы округлые или продолговато-почковидные; покрывальце в виде плёнчатого колпачка при основании соруса, рано опадающее
Cystopteris - Пузырник
Растение с коротким толстым корневищем, образующее воронковидные розетки листьев. Черешок короче листовой пластинки, с редкими светло-бурыми чешуйками. Листовая пластинка в очертании эллиптическая или ланцетная.
Athyrium - Кочедыжник
Растение с длинным горизонтальным корневищем и одиночным расставленными листьями. Черешок длинный, приблизительно равный листовой пластинке, с чёрными или чёрно-бурыми чешуйками.Листовая пластинка в очертании треугольная
Diplazium -Диплазий
Род 1. Athyrium Roth - Кочедыжник В Средней России один вид: 2.A.ftHx-femina(L.) Roth-К. женский, или Женский папоротник.
Род 2. Cystopteris Bernh. - Пузырник
1. Корневище короткое. Листья образуют розетку. Листовая пластинка дважды перисторассечённая с глубоко перистолопастными или раздельными сегментами С. fragilis (L) Bernh, - П. ломкий
Корневище ползучее. Листья одиночные. Листовая пластинка трижды перисторассечённая С. sudetica А.Вг, et Milcfe - П. судетский
Род 3. Diplazium Sw. -Диплазий
В Средней России один вид: 5. D. sibiricum (Turcz. ex G.Kunze) Kurata - Д. сибирский.
СЕМЕЙСТВО DRYOPTERIDACEAE (ASPIDIACEAE) - ЩИТОВНИКОВЫЕ
Насчитывает примерно 25 родов и 1000 видов, произрастающих в лесных и горных районах земного шара. В Средней России 3 рода.
Растение до 25 см высотой, с ползучим корневищем. Листья (вайи) одиночные, в очертании треугольные. Сорусы без покрывальца2. Gymnocarpium - Голокучник
Более крупные растения с коротким корневищем, образующие розетки. Сорусы с покрывальцем... 2
Покрывальце почковидное, цел ьнокрайнее, прикрепляется по радиусу. Листья кожистые или травянистые, черешки их по крайней мере не короче 1/4-1/5 длины пластинки; зубцы сегментов второго порядка могут быть заострёнными, но всегда без длинных щетинок.
Dryoptehs - Щитовник
Покрывальце округлое, зубчатое по краю, щитковидное, прикрепляется в центре. Листья кожистые, с очень короткими черешками, густо усаженными плёнчатыми чешуйками; зубцы сегментов второго порядка оканчиваются длинными щетинками
Polystichum - Многорядник
Род 1. Dryopteris Adans. – Щитовник 1. Листья дважды перисторассечённые, ланцетные или продолговатые
Листья трижды (иногда почти четырежды) перистые, в нижней части значительно более широкие. Спороносные листья кожистые, длиннее стерильных, счерешками, почти равными по длине пластинке.
Нижний сегмент листа яйцевидно- или ланцетно-треугольный, рассечённый на 5-6 пар явственных долей. D. cristata (L.) A.Gray - Щ. гребенчатый
Спороносные листья травянистые, одинаковой длины со стерильными, черешок много короче пластинки. Нижний сегмент листа узколанцетный, рассечённый на 12-15 и более пар явственных долей. D. filix-mas (L.) Schott - Щ. мужской, или Мужской папоротник
Пластинка листа снизу голая, трижды перисторассечённая. Первая нижняя доля нижнего сегмента примерно в 1,5 раза длиннее верхней. Черешок покрыт светло-бурыми плёнками. D. cartusiana (Vill.) H.P.Fuchs - Щ. Картузиуса
Пластинка листа снизу покрыта чешуйками и железистыми волосками, трижды или четырежды перисторассечённая. Первая нижняя доля нижнего сегмента в 2,5 раза и более длиннее верхней.
Черешок покрыт плёнками с продольной тёмной полоской. D. expanse (C.Presi) Fraser-Jenkins et Jermy - Щ. распростёртый
Род 2. Gymnocarpium Newm. - Голокучник
Конечный се'гмент листа примерно равен боковым. Листья светло-зелёные, голые. G. dryopteris (L) Newm. - Г. Обыкновенный.
Конечный сегмент листа заметно крупнее боковых. Листья тёмно-зелёные, снизу с железистым опушением robertianum (Hoffm.) Newm. - Г. Робертов.
Род 3. Polystichum Roth - Многорядник
В Средней России один вид: P. braunii (Spenn.) Fee - М. Брауна.
СЕМЕЙСТВО THELYPTERIDACEAE - ТЕЛИПТЕРИСОВЫЕ
Насчитывает примерно 15 родов и 1100 видов, произрастающих не по всему земному шару. В Средней России 2 рода.
1. Нижняя пара сегментов листа отклонена книзу, начиная со второй пары, сегменты сросшиеся основаниями. Листья снизу опушённые. Черешок и стержень листа с буроватыми чешуйками
Phegopteris - Фегоптерис
Нижняя пара сегментов листа не отклонена, сегменты не сросшиеся. Листья голые. Черешок и стержень листа без чешуек.
Thelypteris - Телиптерис
Род 1. Phegopteris Fee - Фегоптерис
В Средней России один вид: 13. Ph. connectilis (Michx.) Watt - Ф. связывающий.
Род 2. Thelypteris Schmidel - Телиптерис В Средней России один вид: Т. palustris Schott - Т. болотный.
СЕМЕЙСТВО ASPLENIACEAE - КОСТЕНЦОВЫЕ, или АСПЛЕНИЕВЫЕ
Насчитывает примерно 10 родов и около 700 видов, распространённых по всему земному шару. В Средней России один род.
Род Asptenium L - Костенец
Листья перистые, с короткими черешками и многочисленными округлыми сегментами
Листья дважды или трижды перисторассечённые или 2-3-раздельные, с длинными черешками
Черешок и стержень листа чёрные или бурые по всей длине, блестящие. A. trichomanes L - К. волосовидный
Черешок и стержень листа зелёные, по крайней мере в верхней части. A. viride Huds. - К. зелёный
Листья дважды-трижды перистораздельные, сегменты их широкие.
Л. ruta-muraria L. - К. постенный
Листьядвух-трёхраздельные, сегменты их линейные A, septentrionale L) Hoffm. - К. северный
СЕМЕЙСТВО HYPOLEPIDACEAE - ГИПОЛЕПИСОВЫЕ, или ОРЛЯКОВЫЕ
Насчитывает 6 родов и около 100 видов, распространённых преимущественно в тропических и субтропических областях Восточного полушария. В Средней России только один род Pteridium Gled. ex Scop. -Орляк который представлен одним видом: P. aquilinum (L.) Kuhn ex Decken - О. обыкновенный.
СЕМЕЙСТВО POLYPODIACEAE - МНОГОНОЖКОВЫЕ
Насчитывает около 30 родов и 1000 видов, распространённых по всему земному шару. В Средней России только один родPolypodiumL. -Многоножкас одним видом: P. vulgare L. -М. обыкновенная.
СЕМЕЙСТВО OPHIOGLOSSACEAE - УЖОВНИКОВЫЕ
Насчитывает 3 рода и более 80 видов, распространённых преимущественно в Северном полушарии. В Средней России 2 рода.
Стерильная часть листа цельная, спорангии в виде линейного двурядного колоса Ophioglossum- Ужовник
Стерильная часть листа перисторассечённая, спороносная часть метельчато разветвлённая Botrychium – Гроздовник.
Род 1. Botrychium Sw. - Гроздовник
Стерильная часть листа в очертании продолговатая, просто перисторассечённая, с цельными сегментами, почти сидячая, отходит примерно от середины листа S. tunaria (L.) Sw. - Г. Полулунный.
Стерильная часть листа в очертании треугольная или яйцевидная, дважды-трижды перистая, отходит либо от основания, либо выше середины листа
Стерильная часть листа с длинным черешком отходит от основания листа S. multifidum (S.G.Gmel.) Rupr. - Г. Многораздельный.
Стерильная часть листа сидячая или с коротким черешком, отходит выше середины листа.
Стерильная частьлиста широкотреугольная, трижды перистая, сидячая S. virginianum (L.) Sw. - Г. Виргинский.
Стерильная частьлиста треугольно-продолговатая или яйцевидная, дважды перистая, сукороченными
и обычно сливающимися верхними перьями В. matricariifolium A.Braun ex Koch - Г. Ромашколистный.
Род 2. Ophioglossum L. - Ужовник
В Средней России встречается один вид: 25.0. vulgatum L. -У. обыкновенный.
СЕМЕЙСТВО SALVINIАСЕАЕ - САЛЬВИНИЕВЫЕ
Включает один род Salvinia Seguier- Сальвиния с 12 видами, распространёнными преимущественно в Северном полушарии. В Средней России только один вид: S.natans ( L.) АИ.- С. Плавающая.
Глава 2. ХАРАКТЕРИСТИКА ВИДОВ ОТДЕЛА ПАПОРОТНИКООБРАЗНЫЕ ПЕНЗЕНСКОЙ ОБЛАСТИ
2.1 Семейство Dryopteridaceae – Щитовник
Крупный, 40-100(120) см высотой, многолетний папоротник с утолщённым корневищем и бокальчато расположенными тёмно-зелёными ланцетными или продолговатыми дважды перисторассечёнными листьями с короткими черешками. Черешок и стержень листа вплоть до самой верхушки обычно густо покрыты светло-коричневыми плёнчатыми чешуйкам и. Сегменты пластинки первого порядка линейно-ланцетные, глубоко пери-сторассечённые, стойкой оттянутой верхушкой, с продолговатыми тупыми и тупо-зубчатыми сегментами второго порядка. Сорусы расположены на нижней поверхности конечных сегментов листьев и покрыты почковидным покрывальцем. Споры созревают в июне-августе.
Распространён в Европе, на Кавказе, в Средней Азии, Северной Америке. В России встречается повсеместно в лесных районах европейской части, на юге Сибири (Алтай, Саяны).
Растёт в тенистых широколиственных и хвойно-широколиственных лесах на богатой умеренно влажной почве; на севере часто связан с известняками и каменистыми почвами; часто встречается в большом числе экземпляров, хотя сплошных зарослей не образует.
Декоративное растение, выведено несколько садовых форм. Используется в медицине и ветеринарии как противоглистное средство. Растение ядовито, поэтому самолечение опасно для жизни. Корневище используется также для дубления кож и окраски их в жёлтый цвет.
Многолетнее растение с утолщённым и коротким корневищем. Листья 30-50 см длиной, триады перисто-рассечённые, с длинными черешками, одетыми одноцветными светло-бурыми овальными плёнчатыми чешуйками. Форма пластинки варьирует - от продолговатой до треугольно-яйцевидной. Первая нижняя доля бокового сегмента не более чем в 1,5 раза длиннее верхней; первые его доли супротивные. Конечные зубчики листа заканчиваются шиловидным острием. Спороносит в июле-августе.
Распространён в Европе, на Кавказе, в восточной части Северной Америки. В России не заходит восточнее Енисея и отсутствует в степных районах.
Растёт в разнообразных лесах, преимущественно темнохвойных, часто у стволов ели, между корневыми лапами и у разлагающихся пней. В лесной зоне - самый обыкновенный из папоротников.
Многолетнее растение 30-50 см высотой, с толстоватым укороченным корневищем. Листья тёмно-зелёные, кожистые, лоснящиеся, с длинными черешками, дважды перисторассечённые. Спороносные листья удлинённо-ланцетные, торчащие, достигают в длину в отдельных случаях до 70 см; вегетативные листья более короткие, продолговатые. Сегменты первого порядка на вегетативных листьях сближенные, а на спороносных - несколько расставленные. Сорусов немного, они расположены в два ряда. Спороносит в июле-сентябре.
Распространён в северной и средней частях Европы и в Северной Америке. В России встречается в лесных районах европейской части и в Западной Сибири. 8 Средней России во всех областях, но к югу реже.
Растёт на моховых и травяных болотах, в заболоченных лесах и кустарниках; чаще всего на кочках или приствольных повышениях. Нередко, но спорадически.
2.2 Семейство Thelypteridaceae – Телиптерисовые
Многолетнее растение 30-60 см высотой стойким чёрным ползучим корневищем, листовые пластинки желтовато-зелёные, на длинных черешках, продолговатые или ланцетные, дважды перистые; сегменты их ланцетно-линейные, уменьшающиеся в размерах к основанию пластинки, глубоко перисто-раздельные с продолговатыми долями; у спороносящих листьев они треугольно-серповидные благодаря закручивающимся внутрь краям, прикрывающим сорусы. Споры созревают в июле-сентябре.
Распространён в Европе, на Кавказе, в Средней Азии и Северной Америке. В России - по всей территории в лесной полосе.
Образует обширные заросли на мокрых заболоченных лугах и в заболоченных лесах, преимущественно в черно- ольшаниках, а также по краям зарастающих торфяных водоёмов, часто образуя сплавину.
Многолетний папоротник 15-30 см высотой, с тонким ползучим корневищем. Листовые пластинки тёмно-зелёные, стреловидно-дельтовидные, с оттянуто-заострённой верхушкой, дважды перисторассечённые, с удлинёнными черешками, превышающими по длине пластинки. Сегменты ланцетные или линейно-ланцетные, заострённые, глубоко перисто-раздельные с продолговатыми тупыми цельнокрайними долями; нижняя пара их резко отклонена книзу. Споры созревают в июне-августе.
Распространён по всей Европе, на Кавказе, в Северной Америке. В России встречается практически по всей лесной территории. В Средней России обычен в северных областях, к югу редеет.
Растёт в тенистых широколиственных и хвойных лесах, ольшаниках на достаточно богатой умеренно увлажнённой почве, часто в большом числе экземпляров.
2.3 Семейство Hypolepidaceae - Орляковые
Один из наиболее крупных, 30-150 см высотой, и обычных папоротников с мощно развитой, сильно разветвлённой системой чёрных горизонтальных и вертикальных корневищ. Листовые пластинки плотные, в очертании треугольные или овально-дельтовидные, дважды-трижды перистые, с длинными и толстыми черешками; сегменты второго порядка продолговато- или линейно-ланцетные, тупые, иногда при основании лопастные или перисто-надрезанные. Сорусы расположены по прикрывающему их краю пластинки (спорангии развиваются не каждый год!). Споры созревают в июле-августе.
Распространён по всему земному шару, кроме полярных районов, степей и пустынь. Повсеместно встречается и в России. Массовый вид на всей территории Средней России.
Растёт в хвойных, преимущественно сосновых, и лиственных, главным образом берёзовых, лесах на легких и бедных почвах, иногда на известняках; нередко доминируют в травяном покрове.
Легко размножается вегетативным путем, заселяя обширные площади на вырубках и гарях, а также на сенокосах и пастбищах, где считается трудно искореняемым сорняком.
Молодые побеги - распускающиеся листья в стадии «улитки» - на Востоке употребляются в пищу; корневища содержат до 46% крахмала и местами используются для приготовления клея, в пивоварении, на корм свиньям, но для прочего скота ядовиты. Листья имеют своеобразный запах и противогнилостные свойства и используются для завёртывания плодов и овощей, из них получается хорошая подстилка для скота, улучшающая к тому же качество навоза. Зола содержит соли калия и применяется при мыловарении и производстве стекла.
2.4 Семейство Salviniaceae - Сальвиниевые
Водный однолетний разноспоровый папоротник с тонким разветвлённым стеблем, 3-10 см длиной, плавающий на поверхности водоёмов. Листья собраны потри в мутовки: два - надводных, один - подводный. Надводные листья овальные, тупые, при основании слабо сердцевидные, с верхней стороны с щетинистыми белыми волосками, сидящими на небольших бородавках, расположенных косыми рядами; снизу густо опушены бурыми волосками. Подводный лист от самого основания разделён на нитевидные доли. В основании подводных листьев собраны шарообразные сорусы - по 3-8 вместе; в одних сорусах находятся макроспорангии (каждый с одной макроспорой), в других - микроспорангии с большим числом (до 64) микроспор. Споры созревают в августе-сентябре.
Распространена в Средней и Южной Европе, Северной Африке, на Кавказе, в Средней и Центральной Азии, Японии, Китае, Северной Америке. В России встречается преимущественно в южной половине европейской части, в Западной Сибири и на Дальнем Востоке. В Средней России проходит северная граница ареала. Включена в Красную книгу Московской области.
Предпочитает водоёмы со стоячей и медленно текущей, хорошо прогреваемой водой, особенно старицы крупных рек. На северном предепе ареала крайне редка и рассеянна, но в местах экологического оптимума (например, в низовьях Волги) представлена массово.
Размножается и распространяется преимущественно вегетативно: побеги её легко ломаются и дают начало новым растениям; местами способна сплошь заполнить поверхность водоёма. Предполагается, что сальвинию могут разносить водоплавающие птицы.
2.5 Семейство Ophioglossaceae – Ужовниковые
Короткокорневищное многолетнее растение 8-25 см высотой, с одной или двумя вегетативными листовыми пластинками и черешками, отходящими от основания или нижней части общего стержня листа. Пластинка мясистая, тёмно-зелёная, широко-треугольная, до 7 см длиной и 10 см шириной. Доли пластинки первого порядка двоякоперисто-рассечённые, раздельные или надрезанные, конечные дольки - яйцевидные или ромбически-яйцевидные, тупые, слабо городчатые. Спороносная метёлка обычно треугольно-овальная, дважды- или трижды разветвлённая. Споры созревают в июне-июле.
Распространён практически по всему земному шару, за исключением пустынных районов и Африки. В России встречается в лесных районах европейской части, а также в Западной Сибири.
Растёт под пологом леса, на опушках и полянах, на достаточно влажной почве, по окраинам болот и берегам морских заливов; нередко, но спорадически.
Небольшое, 5-20 см высотой, многолетнее растение. Вегетативная часть листа несёт несколько пар почковидно-ромбических или полулуных сегментов, цельнокрайних или тупозубчатых. Спороносная метёлка обычно дважды-трижды перистая, реже просто перистая, вместе с ножкой обычно превышает вегетативную часть. Споры созревают в июне-июле.
Распространён практически по всему земному шару, исключая пустынные районы. В России встречается по всей территории, но спорадически.
Растёт на сыроватых, обычно мшистых луговинах, полянах, опушках, по склонам речных долин и оврагов, иногда на выходах мела и известняка, чаще в небольшом числе экземпляров. 8 зависимости от устойчивости общего экологического фона, в основном гидрологического режима, может длительное время существовать на одном и том же месте или полностью исчезать из покрова в течение 2-3 лет.
2.6 Семейство Onocleaceae - Оноклеевые
Крупный, до 120 см высотой, многолетний папоротник с воронковидно расположенными вегетативными листьями, между которыми в центре находятся несколько спороносных листьев, более коротких, с почти цилиндрическими сегментами, сначала зелёных, а при созревании - бурых. Вегетативные листья продолговатые или обратноланцетные, перисто-рассечённые, с перисто-раздельными и лопастными сегментами и короткими широко желобчатыми черешками; размер сегментов книзу уменьшается; доли второго порядка цельнокрайние. Спороносит в августе-сентябре.
Распространён в районах с умеренным климатом северного полушария, в том числе на всей территории России, но в южных районах встречается реже.
Приурочен к лесным долинам небольших речек и ручьёв, к днищам оврагов, где растёт на наносах почвы, местами образуя обширные заросли.
Декоративное растение. Использовалось в народной медицине.
2.7 Семейство Athyriaceae - Кочедыжниковые
Многолетнее растение до 100 см высотой. Листья крупные, дважды или почти триджы перисторассечённые, со сравнительно короткими черешками, покрытыми буроватыми чешуйками. Сорусы серповидные, реже овальные, плотно расположенные; покрывальце сорусов по краю надрезанное. Спороносит в июле-августе.
Широко распространён в умеренном поясе северного полушария, в том числе почти на всей территории России, за исключением степных районов.
Обычен во влажных лесах различных типов, местами образуя заросли на полянах. Интенсивно распостраняется; споры хорошо прорастают на влажной, лишённой растительного покрова почве, вдоль тропинок, на гнилых пнях и т. п.
Считается ядовитым для лошадей и крупного рогатого скота. Применяется в народной медицине как глистогонное средство
Небольшой, очень изящный и хрупкий папоротник. Корневище укороченное. Листья сидят пучками, 10-30 см длиной, мягкие, светло-зелёные, ажурные. Сегменты первого порядка с черешочками, самые нижние короче соседних; сегменты второго порядка продолговатые, округло-лопастные или зубчатые. Черешок тонкий, ломкий, буроватый. Сорусы расположены однорядно с обеих сторон средней жилки. Спороносит в июне-июле.
Распространён в Северной и Средней Европе, на Кавказе и в Средней Азии. В России встречается повсеместно, но неравномерно, преимущественно в лесной полосе и в горных районах, хотя местами довольно обыкновенен.
Растёт по влажным, часто глубоким лесистым оврагам на местах, лишённых растительного покрова, предпочитая глинистую и, особенно, известковую почву. В оптимальных условиях, например, в старых камнеломнях и на скалистых обнажениях, в массе расселяется по вертикальным стенам известняка.
Считается ядовитым, так как содержит синильную кислоту.
Диплазий сибирский
Многолетнее растение до 50 см высотой, с длинным ползучим корневищем. Близко расположенные друг к другу довольно крупные листья обычно образуют красивую мозаику. Сегменты первого порядка в числе 15-20 пар продолговато-ланцетные, сегменты второго порядка продолговатые, иногда выгрызанно-выемчатые, с городчатыми по краю лопастями. Черешки с черноватыми плёнчатыми чешуями. Сорусы овальные или округлые, выемчатые по краям. Спороносит в июле-августе.
Распространён в Скандинавии и японо-китайском регионе. В России встречается спорадически почти по всей территории таёжной зоны, но преимущественно в Сибири и на Дальнем Востоке. В Средней России свойствен северным нечернозёмным районам; везде редок. Включён в Красную книгу Московской области.
Приурочен к влажным, чаще темнохвойным лесам, лентовидно разрастается вдоль небольших лесных ручьёв или речек; предпочитает известковую почву.
Весьма декоративен.
Невысокий, 10-25 см высотой, многолетний папоротник с сильно разрастающимся длинным разветвлённым корневищем, поэтому один клон может занимать значительную площадь. Листовые пластинки светло-зелёные, в очертании широкотреугольные; доли их на черешках, дважды перисторассеченные, сегменты второго порядка яйцевидные или ланцетные, перисто-раздельные, обычно с тупыми округлыми цельнокраиними конечными дольками. Сорусы удалены друг от друга, без покрывальца. Споры созревают в июне-июле.
Распространён в Европе, на Кавказе, в Средней Азии, Японии, Китае, Северной Америке. В России встречается в лесной полосе почти повсеместно, кроме крайнего севера и Дальнего Востока.
Растёт преимущественно в темнохвоиных (еловых) и смешанных лесах, на небогатой и умеренно влажной почве, часто разрастаясь в просветах крон и иногда на лесосеках - на мелких порубочных древесных остатках. Всюду достаточно обыкновенен.
Заключение
В этой работе я дал общую характеристику видам отдела Папоротниковидные, классификацию папоротников Пензенской области и подробное описание видов которые произрастают на нашей территории. Мной были описаны: ужовник обыкновенный, гроздовник полулунный, гроздовник многораздельный, страусник обыкновенный, кочедыжник женский, диплазий сибирский, цистоптерис ломкий, голокучник обыкновенный, щитовник игольчатый, щитовник гребенчатый, щитовник мужской, телептерис болотный, фигоптерис связывающий, орляк обыкновенный, сальвиния плавающая. Было дано: их особенности, распространение и хозяйственное значение.
Литература
1. Благовещенский В.В. и др. Определитель растений Среднего Поволжья. Л.: Наука, 1984.392 с.
2. Васюков В.М. Конспект флоры сосудистых растений заповедника «Приволжская лесотепь»: Вып.1. Биологическое разнообразие и динамика природных процессов в заповеднике «Приволжская лесостепь». Пенза, 1999. С. 47-80.
3. Губанов И.А., Киселева К.В., Новиков B.C., Тихомиров В.Н. Определитель сосудистых растений центра Европейской России. 2-е изд. доп. и перераб. М.: Аргус, 1995. 560 с.
4. Еленевский А.Г., Соловьева В.Н., Тихомиров В.Н. Ботаника высших, или наземных растений. М.: Академия, 2000. 428 с.
5. Киреев Е.А., Деревянко А.А., Кутарьев В.П. Дополнение к флоре лесного участка «Верховье Суры» ГПЗ «Приволжская лесостепь». //Летопись природы заповедника «Приволжская лесостепь», 2001. Рукопись.
6. Комарницкий Н.А. и др. Ботаника: систематика растений. М.: Просвещение, 1975. 608 с.
7. Красная книга РСФСР. Растения, М.: Россельхозиздат, 1988. 590 с.
8. Маевский В.В. Володушка длиннолистная. /Красная книга Саратовской области. Саратов: Регион. Приволж. изд-во «Детская книга», 1996. С. 32.
9. Маевский П. Ф. Флора Средней полосы Европейской части СССР. /Под.
ред. Б. П. Шишкина. 9-е изд. и доп. Л.: Колос, 1964. 890 с.
10. Новикова Л. А. Растительность Пензенской области: Методические указания к спецкурсу для студентов естественно-географического факультета. Пенза: ПГПУ, 2000. 38 с.
11. И.Силаева Т.Е., Тихомиров В.Н., Майоров СР. Редкие и исчезающие растения Мордовии: Учебное пособие. - Саранск: изд-во Морд, ун-та, 1996.-72 с.
12. Солянов А.А. Основные задачи современной региональной флористики. //Проблемы охраны и рационального использования природных экосистем и биологических ресурсов: Мат. Всерос. науч-практ. конф., посвящ. 125-летию И.И. Спрыгина. Пенза, 1998. С. 64-66.
13. Солянов А.А. Флора Пензенской области, г. Пенза: ПГПУ, 2001. 310 с.
14. Спрыгин И.И. Материалы к флоре губерний Пензенской и Саратовской. //Тр. Общ. естествоиспыт. при Импер. Казанском ун-те. Казань, 1896, т.29, вып.6. С. 1-75.
15. Спрыгин И.И. Почвенные и ботанические исследования в Пензенском и Городищенском уездах в 1896-1899гг. //Тр. Общ. естествоиспыт. при Импер. Казанском ун-те. Казань, 1900, т. ЗЗ, вып.5. С. 1-60.
16. Спрыгин И.И. Сосна и ее спутники в Пензенском уезде. //Тр. Общ. естествоиспыт. при Импер. Казанском ун-те. Казань, 1908, т.61, вып.З. С. 1-157.
17. Спрыгин И.И. О некоторых редких растениях Пензенской губернии. //Тр. ПОЛЕ Пенза, 1914, вып.1. С. 3-17.
18. Спрыгин И.И. О некоторых лесных реликтах Приволжской возвышенности. //Уч. зап. Казанского гос. ун-та, 1936, т.96; кн 6. С. 1-117.
19. Спрыгин И.И. Из области Пензенской лесостепи. Ч.З. Степи песчаные, каменисто-песчаные, солонцеватые, на южных и меловых склонах. Пенза: Гос. ком. по охр. окр. ср. Пенз. обл., 1998. 140 с.
20. Тахтаджан А.Л. Система магнолиофитов. Л.: Наука, 1987. 439 с.
21. Флора СССР. /Под. ред. Б.К. Шишкина М-Л.: АН СССР, т.16 - 1950, т.17 -1951.
22. Цвелев Н.Н. Семейство Зонтичные. //Жизнь растений, т.5 (I). С.302-30.
© 2010 Интернет База Рефератов