
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Курсовая работа: Синаптическая передача. Медиаторы
Курсовая работа: Синаптическая передача. Медиаторы
Синаптическая передача. Медиаторы
Введение
Ч. Шеррингтон обосновал представление об отсутствии межклеточной непрерывности в нервной системе и ввел понятие синапс для обозначения структуры, которая опосредует передачу сигнала от окончаний аксона к эффекторной клетке – нейрон), мышечному волокну, секреторной клетке железы. Синапс состоит из пресинаптического окончания и постсинаптической мембраны, разделенных синоптической щелью, которая заполнена рыхлым коллагеноподобным веществом.
Существуют два способа синаптической передачи – электрический и химический.
Возможно и сочетание обоих механизмов, электрического и химического, в одном смешанном синапсе, однако в нервной системе млекопитающих преобладают чисто химические синапсы.
В электрических синапсах, количество которых в нервной системе относительно невелико, потенциал действия пресинаптических окончаний обеспечивает ток, который деполяризует постсинаптическую мембрану. Морфологическую основу электрической передачи составляет щелевой контакт, для которого характерны тесное прилегание пре- и постсинаптической мембран, большая площадь контакта этих мембран, наличие ультраструктур, снижающих электрическое сопротивление в области контакта, – своего рода каналов, организованных в ввде правильной сети между пре- и постсинаптической мембраной.
Электрофизиологическими критериями электрической синаптической передачи являются: 1) отсутствие синаптической задержки; 2) проведение возбуждения в обоих направлениях; 3) независимость от потенциала пресинаптической мембраны; 4) устойчивость к изменениям концентрации ионов кальция и магния в среде, к асфиксии, низкой температуре, некоторым фармакологическим воздействиям.
Функциональная роль электрических синапсов состоит в осуществлении срочной передачи сигналов, обеспечивающей синхронизацию электрической активности группы нейронов, например группы мотонейронов во время прыжковых движений лягушки или плавательных движений рыбы. Электрические синапсы обнаруживаются между нервными клетками, однотипными по структуре и функциям.
Эволюция нервной системы сопровождается уменьшением числа электрических синапсов в пользу другого способа передачи – химического. В химическом синапсе нервный импульс вызывает освобождение из пресинаптических окончаний химического посредника – нейромедиатора, который диффундирует через синоптическую щель и вступает во взаимодействие с белками-рецепторами постсинаптической мембраны, В результате происходит генерация постсинаптического потенциала.
Химический механизм синаптической передачи по сравнению с электрическим более эффективно обеспечивает основные функции синапса: 1) одностороннее проведение сигнала; 2) усиление сигнала; 3) конвергенцию многих сигналов на одной постсинаптической клетке, пластичность передачи сигналов.
Химические синапсы передают два вида сигналов – возбуждающий и тормозной. В возбуждающих синапсах нейромедиа-тор, освобождаемый из пресинаптических нервных окончаний, вызывает в постсинаптической мембране возбуждающий пост-синаптический потенциал – локальную деполяризацию, а в тормозных синапсах – тормозной постсинаптический потенциал, как правило, – гиперполяризацию. Снижение сопротивления мембраны, происходящее во время тормозного постсинаптического потенциала, ведет к короткому замыканию возбуждающего постсинаптического тока, тем самым ослабляя или блокируя передачу возбуждения.
В настоящей работе рассмотрены организация пресинаптического окончания, механизмы экзоцитозо. Ейромедиаторов, химическая природа и функциональные особенности медиаторов.
1. Морфофункциональная организация химического синапса. Квантовая теория освобождения нейромедиатора
Одним из самых детально изученных химических синапсов считается нервно-мышечное соединение скелетных мышц, в котором нейромедиатором служит ацетилхолин. Относительная доступность этого синапса и удобство для изучения позволили нобелевскому лауреату Б. Катцу в 50‑е годы провести мик-роэлектрофизиологические исследования, которые вместе с данными электронной микроскопии составили основу квантовой теории синаптической передачи, справедливой для самых разных химических синапсов.
Согласно этой теории процесс освобождения нейромедиатора складывается из отдельных элементарных реакций, каждая из которых представляет собой выход одного кванта нейромедиатора. Когда потенциал пресинаптической мембраны находится на уровне покоя, т.е. к пресинаптическим окончаниям не поступают нервные импульсы, кванты нейромедиатора тоже освобождаются, но спонтанно и с низкой скоростью. Ответом постсинаптической мембраны на отдельные кванты является возникновение миниатюрных постсинаптических потенциалов, в случае нервно-мышечного синапса они называются миниатюрными потенциалами концевой пластинки. Деполяризация пресинаптической мембраны во время нервного импульса ведет к практически синхронному освобождению большого количества квантов – до нескольких сотен. В результате возникает вызванный постсинаптический потенциалов нервно-мышечном синапсе он называется потенциалом концевой пластинки), который, в случае достижения пороговой амплитуды, ведет к генерации потенциала действия в постсинаптической клетке.
Освобождение индивидуальных квантов представляет собой статистический процесс: нервное окончание содержит п элементарных единиц, каждая из которых имеет независимую от других единиц вероятность ответа на нервный импульс. Если средняя вероятность ответа равна Ру то среднее число квантов, освобожденных в ответ на нервный импульс, определяется уравнением т=пР.
Повышение вероятности освобождения квантов при деполяризации пресинаптической мембраны связано с открыванием потенциал-зависимых Са+-каналов мембраны и входом Са в соответствии с их электрохимическим градиентом в пресинаптичское окончание. Таким образом, ионы Са участвуют в процессе электросекреторного сопряжения.
Квантовый характер синаптической передачи обусловлен тем, что нейромедиатор хранится в синаптических пузырьках. Присутствие везикул диаметром 40–200 нм, окруженных относительно плотной для электронов мембраной толщиной 4–5 нм, является характерной морфологической особенностью химических синапсов. Объем синаптического пузырька в двигательных нервных окончаниях диафрагмы крысы составляет 5,2–10 нм. Такой пузырек может содержать порядка АЛ О молекул АХ, что вполне соответствует результатам оценки молекулярного состава кванта АХ – -10 молекул.
Синаптические пузырьки в аксоплазме нервного окончания сосредоточены в области, приближенной к пресинаптической мембране, около синаптической щели, причем пузырьки распределяются неравномерно, группируясь у периодически выступающих в аксоплазму утолщений пресинаптической мембраны – активных зон. По-видимому, в активных зонах находятся скопления потенциал-зависимых Са+-каналов, обеспечивающих вход Са+ в пресинаптическое окончание во время потенциала действия. В активных зонах обнаружены регулярно расположенные розеткообразные мембранные частицы диаметром около 15,0 нм, количество которых возрастает во время деполяризации пресинаптической мембраны. Можно предполагать, что эти частицы представляют собой точки слияния синаптических пузырьков с активной зоной, т.е. участки экзоци-тоза нейромедиатора в синаптическую щель.
При физиологических значениях частоты и длительности стимуляции нерва не удается выявить изменения количества синаптических пузырьков, несмотря на наблюдаемые изменения количества освобождаемого медиатора. Только при воздействиях, вызывающих истощение пресинаптических запасов медиатора, отмечается некоторый параллелизм изменений постсинаптических потенциалов и морфологических характеристик пресинаптических окончаний.
В период начального бурного освобождения медиатора, когда резко повышается частота спонтанных миниатюрных постсинаптических потенциалов, в области активных зон появляются многочисленные омегаподобные впячивания пресинаптической мембраны, которые соответствуют слипанию мембраны пузырька с участком освобождения. В фазу снижения квантового состава вызванных постсинаптических потенциалов наблюдается уменьшение количества синаптических пузырьков и увеличение площади поверхности пресинаптического окончания за счет встраивания мембран синаптических пузырьков в пресинаптическую мембрану. В дальнейшем идет процесс рециклизации синаптических пузырьков: от пресинаптической мембраны в нервное окончание отпочковываются мембранные структуры, которые сливаются, образуя окруженные мембраной цистерны; затем от цистерн отделяются синаптические пузырьки, вновь заполняемые медиатором, синтезированном в цитоплазме.
Процесс рециклизации синаптических пузырьков прослежен с помошью пероксидазы хрена. При добавлении этого маркера в среду инкубации электрическое раздражение нерва сопровождается усиленным захватом маркера синаптическими пузырьками и цистернами. Если после этого перенести препарат в среду, не содержащую пероксидазу хрена, и вновь подвергнуть его стимуляции, то пероксидаза освобождается в среду при экзоцитозе содержимого синаптических пузырьков.
Оценку динамики изменений запасов медиатора в пресинаптическом окончании во время ритмического раздражения нерва можно производить электрофизиологически путем измерения амплитуды и расчета квантового состава постсинаптических потенциалов. Классическими объектами для таких исследований стали нервно-мышечный синапс и симпатический ганглий. Структуры центральной нервной системы, где на теле или дендрите одного нейрона конвергируют много взаимодействующих синапсов, непригодны для таких оценок. Ритмическое раздражение двигательного нерва скелетной мышцы млекопитающего сопровождается быстрым снижением количества освобождаемого в ответ на каждый импульс медиатора, вплоть до некоторого относительно постоянного уровня. Это явление, называемое депрессией, показывает, что запас медиатора, способного к освобождению, ограничен и пополняется медленнее, чем расходуется. Депо АХ в нервно-мышечном синапсе оценивают примерной цифрой 210 квантов. Быстрая депрессия потенциалов концевой пластинки отражает расходование небольшой части депо АХ – так называемой фракции доступного медиатора, которая составляет примерно 10 квантов для нервно-мышечного синапса лягушки.
Интересной особенностью метаболизма пресинаптических окончаний является предпочтительная секреция вновь синтезированного в цитоплазме медиатора. После инкубации нервно-мышечного препарата или симпатического ганглия в среде с меченым предшественником АХ-холином ритмическое раздражение нерва приводило к секреции – АХ. Очевидно, существует фракция расположенных у активных зон синаптических пузырьков, которые после экзоцитоза их содержимого сразу же подвергаются рециклизации, заполняются только что синтезированным в цитоплазме медиатором и вновь готовы к экзоцитозу.
Следует отметить, что наряду с зависимой от ионов Са квантовой секрецией медиатора, которая обеспечивает передачу сигнала через синапс к постсинаптической клетке, а также происходит спонтанно, в отсутствие нервных импульсов, существует постоянная неквантовая утечка молекул медиатора из нервного окончания. В нервно-мышечном синапсе лягушки и млекопитающих неквантовая утечка создает концентрацию АХ в синаптической щели порядка 10~-10~М. Общее количество АХ, секретируемого неквантовым способом, превышает выход АХ, обусловленный спонтанной квантовой секрецией. Предполагается, что неквантовая секреция медиатора играет трофическую роль.
Нервно-мышечный синапс является экспериментальным объектом, удобным для исследования электрофизиологическими методами. Однако изоляция нервных окончаний и синаптических пузырьков для биохимического исследования крайне затруднена вследствие того, что двигательные нервные окончания составляют слишком малую долю от объема ткани скелетной мышцы. Гораздо более адекватным препаратом для изучения экзоцитоза нейромедиаторов являются синаптосомы – пресинаптические окончания, выделяемые из нервной ткани, как правило, вместе с веществом, заполняющим синаптическую щель, и участком постсинаптической мембраны. Деполяризация синаптосом физиологическим раствором с высокой концентрацией К+ вызывает зависимую от Са»^ секрецию нейромедиаторов.
2. Экзоцитоз нейромедиаторов
В нейрохимическом плане лучше других синапсов изучен электромоторный синапс электрического органа рыб, где нейро-медиатором служит АХ. В начале 70‑х годов в лаборатории В. Уит-такера в ФРГ впервые удалось выделить изолированную фракцию синаптических пузырьков из электрического органа ската Torpedo marmorata. Именно на этом объекте с помощью биохимических, иммуноцитохимических методов и ядерного магнитного резонанса получены фундаментальные сведения о структуре и функциях синаптических пузырьков и разработана схема их жизненного цикла.

В аппарате Гольджи сомы нейрона формируются мембранные образования в виде пузырьков, не заполненных медиатором. Эти пузырьки направляются в пресинаптическое окончание с помощью системы быстрого аксонного транспорта. В пресинаптическом окончании пузырьки заполняются медиаторами посредством АТФ-зависимого протонного насоса. Молекулы протонной АТФазы входят в состав мембраны синаптических пузырьков и поддерживают определенный уровень мембранного потенциала. Мембрана синаптического пузырька содержит также стимулируемую кальмодулином Са+-АТФазу, которая обеспечивает поглощение пузырьками ионов Са. Популяция зрелых пузырьков. Циклы экзоцитоз – эндоцитоз повторяются.
Эта схема согласуется с электрофизиологическими данными о квантовом характере секреции нейромедиатора и о численности квантов разных размеров в одном и том же пресинаптическом окончании, а также с радиохимическими сведениями о предпочтительном освобождении вновь синтезированного медиатора. Таким образом, пресинаптическос окончание можно рассматривать как систему, в определенной мере автономную по отношению к телу нейрона.
Синаптические пузырьки диаметром 50–60 нм, так называемые малые прозрачные синаптические пузырьки, аналогичные холинергическим синаптическим пузырькам из электрического организма ската, выделены из разных отделов нервной системы представителей практически всех таксономических групп животных. Эти пузырьки отличаются низкой электронной плотностью содержимого. Они заполнены низкомолекулярными нейромедиаторами в отличие от больших электронно-плотных пузырьков, заполненных медиаторами пептидной природы.
Ключевую проблему в изучении экзоцитоза нейромедиаторов представляет вопрос о механизмах сближения синаптического пузырька с активной зоной пресинаптической мембраны и взаимодействия мембраны пузырька с активной зоной. Имеются данные о том, что эти процессы зависят. от Са+ – универсального внутриклеточного посредника, участие которого в секреторных процессах может быть обусловлено активацией актомиозино-вых филаментов цитоскелета, мембранной фосфолипазы А2, аде-нилатциклазы, Са+/кальмодулин-зависимых протеинкиназ и ряда других Са+-связывающих белков.
Известно, что связывание Са+ с кальмодулином индуцирует фосфорилирование ряда белков синаптосом. Ингибиторы кальмодулина и кальмодулинкиназы блокируют освобождение нейромедиаторов, вызываемое деполяризацией синаптосом. При воздействии различных факторов, влияющих на количество освобождаемого медиатора и на фосфорилирование белков, выявлена корреляция между изменениями секреции нейромедиаторов и фосфорилированием белков синаптосом.
Независимо от того, из какого отдела нервной системы они получены и какой нейромедиатор содержат, малые синаптические пузырьки характеризуются специфическим набором интегральных мембранных белков, к которым относятся синапсины – фосфопротеины, фосфорилируемые цАМФ- и Са+-зависимыми протеинкиназами, синаптофизин гликопротеин, пронизывающий мембрану; синаптобревин – негликозилированный белок, находящийся на цитоплазматической поверхности пузырьков; белок SNAP‑25; синтаксин, синаптогамин, синаптопорин и др.
Особая роль в сближении синаптического пузырька с активной зоной отводится синапсину. Этот белок, который состоит из двух полипептидов с молекулярной массой 86 и 80 кД, ассоциирован с цитоплазматической поверхностью мембраны синаптического пузырька. При микроинъекции фосфорилированной формы синапсинов в пресинаптическое окончание гигантского аксона кальмара наблюдается повышение амплитуды и скорости нарастания постсинаптического потенциала, что свидетельствует об увеличении секреции медиатора; дефосфорилированные формы синапсиноз не вызывали такого эффекта.
Аналогичное увеличение секреции медиатора происходит при микроинъекции Са+/кальмодулина. Показана способность очищенных синапсинов взаимодействовать в зависимости от состояния фосфорилирования с белками мембраны синаптического пузырька и с F‑актином цитоскелета пресинаптического окончания. Предложена следующая схема участия синапсинов в экзоцитозе. В отсутствии деполяризации пресинаптического окончания, когда концентрация Са+ в цитоплазме низка, дефосфорилированный синапсин, связанный с цитоплазматической поверхностью пузырька, взаимодействует с цитоскелетом, обеспечивая резервирование и иммобилизацию пузырька. При деполяризации пресинаптической мембраны происходит вход Са+ в пресинаптическое окончание - активация Са+/кальмодулинкиназы -> фосфорилирование синапсина I -» ослабление связи между синапсином и пузырьком, а также синапсином и F‑актином. В результате синаптический пузырек перемещается вдоль микротрубочек на стратегическую позицию v активной зоны.
Далее наступает цепь реакций, обеспечивающих контакт пузырька с пресинаптической мембраной и его плавление. Здесь опять-таки процесс инициируется Са+, который связывается с другим белком пузырька синаптогамином Именно Са+-синаптогамин взаимодействует с фосфолипидами и с комплексом других белков, регулирующих плавление везикулы, – синаптобревином, синтаксыном и синаптофизином. В заключение происходит активация белка синаптопорина, формирующего пору, какал, через который изливается содержимое везикулы.
В поисках молекулярных механизмов слияния мембраны синаптического пузырька с пресинаптической мембраной выявлено, что ботулинический и столбнячный токсины, блокирующий экзоцитоз нейромедиаторов из синаптических пузырьков, повреждают именно указанную выше триаду белков пузырька – синаптобревин, синтаксин и SNAP‑25.
Ряд других деталей конечного этапа экзоцитоза пока не выяснен. Существует предположение, что выброс нейромедиаторов происходит при активном сокращении стенок пузырька с участием актомиозинподобных белков, активируемых ионами Са.
Синаптическая пластичность означает способность синапсов к функциональным и морфологическим перестройкам в процессе синаптической активности. Свойство пластичности синапсов составляет основу таких явлений, как обучение, память.
Изменение эффективности синапса после активации определяется увеличением или уменьшением амплитуды постсинаптических потенциалов, которые в свою очередь связаны с изменением количества нейромедиатора, высвобождаемого из пресинаптических окончаний.
Существует несколько основных фаз постактивационных изменений синаптической эффективности.
1. Облегчение повышение амплитуды постсинаптического потенциала в начальный период ритмической серии пресинаптических импульсов.
2. Кратковременная посттетаническая потенциация – повышение амплитуды постсинаптического потенциала при ритмической активации нервного окончания в течение десятков секунд.
3. Постактивационная депрессия постсинаптических потенциалов, которая развивается параллельно и взаимодействует с процессом потенциации.
4. Длительная потенциация постсинаптических потенциалов, которая медленно развивается вслед за кратковременной потенциацией и продолжается в течение часов и даже дней.
Эти явления связаны с изменением концентрации Са+ в пресинаптических окончаниях во время ритмической активности. Длительные модификации синаптической эффективности ассоциируются с изменениями фосфорилирования синаптических белков.
Следует упомянуть, что наряду с пресинаптическими механизмами существуют и постсинаптические механизмы, основанные на изменениях чувствительности рецепторов нейромедиаторов.
3. Медиаторы
Развитие представлений о химической медиации нервных импульсов началось в начале века в результате открытий О. Лёви, Дж. Эллиота, ПДейла, которые показали, что передача сигнала в нейроэффекторных соединениях опосредуется высвобождением АХ или норадреналина из нервных окончаний.
До 50‑х годов к медиаторам относили две группы низкомолекулярных соединений, которые сейчас называются «классическими», «традиционными» медиаторами, – амины и аминокислоты. В 60‑е годы Дж. Бэрнсток открыл третью группу медиаторов – пуриновые нуклеотиды. В 1953 г. Ф-Лембек вьщвинул предположение о медиаторной роли пептида – вещества Р, обнаруженного еще в 30‑е годы в мозге и в стенках кишечника в виде вещества, которое усиливало сокращения изолированной кишки и вызывало временную гипотензию. Свое название вещество Р получило от слова «powder», поскольку его первооткрыватели работали с высушенными в виде порошка экстрактами тканей. Разработка иммуноцитохи-мических и радиоиммунологических методов позволила в 70 – 80‑е годы выявить в разных отделах нервной системы позвоночных и безпозвоночных множество пептидов, участвующих в синаптической передаче. Нейропептиды составляют четвертую, самую многочисленную группу медиаторов и, кроме того, выступают как модуляторы действия других медиаторов.
3.1. Принцип Дейла
В 30‑е годы Г. Дейл пришел к выводу, что идентификация медиатора в периферических окончаниях сенсорного нейрона позволяет судить о природе химической передачи в центральном синапсе этого нейрона. Со временем принцип Дейла стал толковаться как постулат, согласно которому каждый нейрон содержит единственное медиаторное вещество, которое высвобождается во всех окончаниях этого нейрона. В таком понимании принцип Дейла, безусловно, не соответствует действительности.
С современных позиций принцип Дейла соответствует, по-видимому, формулировать как положение о метаболической за-еисимости аксона и его окончаний от тела клетки. Известный нейробиолог Дж. Экклс считает, что именно такой смысл вкладывал в свой вывод и сам Г. Дейл.
3.2. Многообразие синаптических медиаторных функций
В настоящее время представление о химическом кодировании сигналов в нервной системе основываются на принципе множественности химических сигналов: в индивидуальном нейроне синтезируется более одного медиатора; каждое пресинаптическое окончание способно высвобождать несколько медиаторов, сочетание которых может не быть одинаковым для разных синапсов одного и того же нейрона.
Ставшее привычным понятие «эргичности» нейрона и синапса можно принимать лишь как условное. Термины «холинергический», «пуринергический», «пептидергический» и т.д. целесообразно употреблять только в случае присутствия в данном нейроне и высвобождении в синапсе конкретного медиатора, не исключая при этом существования других медиаторных веществ и не подразумевая приоритетной роли какого-то одного медиатора по отношению к другим.
Существует деление механизмов преобразования химического сигнала, а соответственно разделение рецепторов медиаторов на две категории – ионотропные и метаботропные. Ионотропные рецепторы составляют единый комплекс с ионофором, так что вызываемое медиатором изменение конформации рецептора ведет к открыванию ионных каналов и быстрым значительным сдвигам проводимости постсинаптической мембраны. Примером являются рецепторы ГАМК, глицина, а также АХ при его взаимодействии с никотиновыми холинорецепторами и часть рецепторов глутамата, аспартата и пуринов.
Метаботропные рецепторы осуществляют постсинаптический эффект путем активации специфических мембранных ферментов, обеспечивающих образование в мембране или в цитозоле постсинаптической клетки вторичных посредников, которые, в свою очередь, специфически активируют определенные ферменты; при этом запускаются каскады ферментативных процессов, ведущих в конечном счете к ковалентной модификации мембранных или цитоплазматических белков. Такой тип действия реализуется гораздо медленнее, чем ионотропный, и сопровождается относительно небольшими сдвигами проводимости иостсинаптическое мембраны. К метаботропной категории относится взаимодействие АХ с мускариновыми рецепторами, постсинаптическое действие катехоламинов и серотонина, части глутаматных рецепторов. Нейропептиды также являются метаботропными медиаторами.
Открытие медиаторов пептидной природы существенно расширило представления о химической медиации сигналов в нервной системе. Совсем недавно классическим образцом химического синапса считалось нервно-мышечное соединение, морфофункциональная организация которого обеспечивает быструю, точно направленную передачу сигнала по «анатомическому» адресу. В системах с «химическим» адресом специфичность передачи сигнала обусловлена не локальной анатомической связью пре- и постсинаптической структуры, а наличием специализированных рецепторов к данному медиатору только на клетках-мишенях, причем такой тип передачи сигнала может быть медленным, диффузным. Именно в передаче такого типа участвуют многие нейропептиды с некоторыми классическими нейромедиаторами, в частности моноаминами, которые тоже могут высвобождаться дистантно по отношению к клетке-мишени. Такое понимание медиаторной функции вплотную приближается к представлению о нейрогормонах, секретируемых в межклеточную жидкость, спинномозговую жидкость или кровь и модулирующих состояние клетки-мишени, расположенной на расстоянии от секретируемой клетки.
Медиаторные вещества делятся на две большие группы: нейромедиаторы, которые осуществляют передачу сигнала в синапсе, и нейромодуляторы, которые регулируют ^передачу сигнала.
3.3. Нейромедиаторная функция
Нейромедиаторная роль вещества в синапсе оценивается следующими критериями.
1. Присутствие медиатора в постсинаптинеском нейроне и, как правилд, неравномерное распределение медиатора в нервной системе. В пресинаптическом нейроне должны находиться молекулы – предшественники медиатора, ферменты его систеза или система специфического транспорта. В синапсе должны быть специфические участки связывания медиатора. Критерий проверяется анатомическими, биохимическими, гистохимическимим методами.
2. Высвобождение медиатора в ответ на деполяризующие стимулы из пресинаптических окончаний посредством Са+-зависимого экзоцитоза. Критерий проверяется физиологическими методами.
3. Идентичность эффектов предполагаемого медиатора и эндогенного нейромедиатора на клетке-мишени; аппликация экзогенного вещества на постсинаптическую клетку должна вызывать такой же эффект, как и физиологическая стимуляция. Взаимодействие медиатора с постсинаптическими рецепторами должно индуцировать сдвиги мембранной проводимости, ведущие к генерации возбуждающих или тормозных постсинаптических потенциалов. Эффекты, вызываемые аппликацией экзогенного вещества или физиологической стимуляцией, должны иметь одинаковые фармакологические характеристики, т.е. подвергаться аналогичным изменениям при действии фармакологичесих средств. Критерий проверяется физиологическими и фармакологическимим методами.
4. Удаление медиатора из области синапса. В синаптической области должны присутствовать специализированные системы инактивации секретированного медиатора, позволяющие завершить его эффект – ферменты деградации, система обратного поглощения пресинаптическим нейроном. Критерий проверяется биохимическими и гистохимическимим методами.
Нужно подчеркнуть, что тестирование типа медиаторной функции по перечисленным критериям представляет собой методически сложную задачу. Особенно это касается критериев и, что обусловлено трудностями доступа к индивидуальным синапсам в ЦНС и ограниченностью существующего набора избирательных фармакологических средств. Определенные успехи обеспечивает применение новых методов – иммуногистомии, рекомбинантных ДНК, клеточных культур.
Итак, нейромедиатор – это вещество, которое синтезируется в нейроне, содержится в пресинаптических окончаниях, высвобождается в синаптическую щель в ответ на нервный импульс и действует на специализированные рецепторные участки постсинаптической клетки, вызывая изменения мембранного потенциала и метаболизма клетки.
3.4 Нейромодуляторы
Понятие «модуляторные вещества», предложенное в 60‑годы Э. Флори, исходит от эндокринологии, от представлений о характере действия гормонов. В современном понимании нейромодуляторы по сравнению с нейромедиаторами имеют следующие характеристики.
1. Нейромодуляторьг не обладают самостоятельным физиологическим действием, а модифицируют эффект нейромедиаторов.
2. Действие нейромодуляторов имеет тонический характер медленное развитие и большую продолжительность действия.
3. Нейромодуляторы не обязательно имеют синаптическое или даже нейронное происхождение. Они могут высвобождаться, например, из глии.
4. Действие нейромодуляторов не сопряжено по времени с эффектом нейромедиатора и не обязательно инициируется нервными импульсами.
5. Мишенью нейромодуляторов может быть не только пост-синаптическая мембрана и не только мембранные рецепторы; нейромодулятор действует на разные участки нейрона, причем его действие может быть и внутриклеточным.
Таким образом, термин «нейромодулятор» является гораздо более широким понятием по сравнению с термином «нейромедиатор».
Различают два основных вида нейромодуляции – пресинаптическая и постсинаптическая.
Пресинаптическая модуляция. Процесс высвобождения многих нейромедиаторов модулируется посредством ауторегуляции: высвобождаемый нейромедиатор воздействует на собственные пресинаптичесие ауторецепторы, уменьшая последующее высвобождение или увеличивая высвобождение. В этой ситуации нейромедиатор одновременно осуществляет и функцию нейромодулятора. Так, например, пресинаптические ос2-адренорецепторы симпатических нервных окончаний опосредуют торможение секреции норадреналина. Пресинаптические ауторецепторы сопряжены с системой аденилатциклазы. По своим фармакологическим характеристикам пресинаптические ауторецепторы обычно отличаются от постсинаптических рецепторов того же нейромедиатора. Известны пресинаптические ауторецепторы глутамата, серотонина, дофамина, ГАМК, гистамина, адренорецепторы, мускариновые холинорецепторы.
Кроме того, существуют пресинаптические гетерорецепторы, которые чувствительны к медиаторам, высвобождающимся из других нейронов. Примером могут служить пресинаптические мускариновые холинорецепторы норадренергических окончаний симпатических нервов, которые взаимодействуют с АХ, секретируюшимся из парасимпатические холинергических аксонов. В этом случае регуляция бывает межнейронной.
Модуляция может происходить на уровне изменений возбудимости нервных окончаний, биосинтеза нейромедиаторов, входа Са+ в нервное окончание и на других этапах экзоцитоза.
Постсинаптическая модуляция. Постсинаптическая модуляция может иметь характер ауторегуляции, когда изменяется активность рецепторов за счет модификации их аффинности или количества, а также вследствие изменений сопряженных с рецепторами систем внутриклеточных и внутримембранных посредников. Примером является десенситизация рецепторов при длительном воздействии нейромедиатора и гиперсенситизация при недостаточности воздействия нейромедиатора.
Постсинаптические рецепторы подвергаются также гетерорегуляции в результате воздействия нейромодуляторных веществ. Значительный интерес вызывает постсинаптическое межрецепторное взаимодействие между сопутствующими медиаторами, прежде всего – нейропептидами и классическими нейромедиаторами.
3.5 Сопутствующие медиаторы
Сопутствующие, или сосуществующие, медиаторы – это синоптические посредники, которые характеризуются прежде всего совместной локализацией и совместным высвобождением. Под совместной локализацией имеется в виду синтез и депонирование медиаторов в одном и том же нейроне, их присутствие в одних и тех же пресинаптических окончаниях, но не обязательно в одних и тех же синаптических пузырьках. Так, низкомолекулярные классические нейромедиаторы депонируются преимущественно в мелких оптически прозрачных пузырьках, а пептидные медиаторы в крупных оптически плотных пузырьках, хотя имеются данные и о случаях локализации этих двух видов медиаторов в одних и тех же оптически плотных пузырьках. Различие в системах депонирования этих двух видах медиаторов обусловлено различиями мест их синтеза: классические нейромедиаторы синтезируются в цитоплазме пресинаптических окончаний и затем поступают в синаптические пузырьки, а пептидные медиаторы синтезируются в аппарате Гольджи, т.е. в соме нейрона, и доставляются в нервные окончания уже упакованными в пузырьки.
Под совместным высвобождением понимается экзоиитоз двух медиаторов в результате одного и того же процесса активации пресинаптического окончания, хотя под процессом активации в данном случае подразумевается не одиночный пресинаптический потенциал действия, а разряд потенциалов действия с той или иной частотой. Еще один признак сопутствующих медиаторов состоит в том, что они вызывают функциональные изменения в одной и той же клетке-мишени.
Иммуноцитохимическими методами в центральных и периферических нейронах прослежены разнообразные виды сочетаний представителей медиаторных групп: 1) несколько классических нейромедиаторов; 2) классический) нейромедиатор + нейропетид несколько нейропептидов, имеющих общую молекулу-предшественник; 4) несколько нейропептидов, кодируемых разными генами. К этим сочетаниям могут добавляться пурины.
3.6 Локализация нейромедиаторных путей
Прежде чем рассматривать наиболее изученные индивидуальные синаптические системы и соответствующие медиаторы, целесообразно рассмотреть общую картину локализации нейромедиаторных путей в мозге, отметив одновременно самые общие характеристики функций соответствующих систем.
Химические синапсы распределены в нервной ткани не в случайном порядке, а организованы в определенных группах нейронов. Для того чтобы уверенно картировать какие-либо меди аторные пути в головной мозге, необходимо иметь доступные и надежные специфические маркеры, с помощью которых можно визуализировать интересующие исследователя межклеточные взаимодействия.
Существуют три основных методических подхода для решения этой задачи. Первый способ – избирательное окрашивание нейронов, выделяющих определенный нейромедиатор, может осуществляться с помощью преобразования естественного медиатора в его флуоресцирующее производное. В этом случае флуоресценция определенных групп клеток поможет выявить специфические связи в структурах мозга. Второй экспериментальный подход связан с введением молекул медиатора, предварительно меченного радиоактивным изотопом. Нейронные окончания, содержащие исследуемый медиатор, способны избирательно захватывать метку. Затем их легко выявить методом авторадиографии. Третий способ обнаружения специфических связей в нервной системе состоит в использовании высоко специфичной способности узнавать либо антигенные детерминанты медиатора, либо определенные ферментные белки, участвующие в метаболизме нейромедиаторов, либо нейрорецептор-ные компоненты на мембране клетки. Последние считаются наиболее убедительным свидетельством в пользу существования конкретных нейрохимических взаимодействий между клетками и зонами мозга. Обычно для иммунохимической идентификации используют флуоресцентный краситель или изотоп, который маркирует антитела. В последние годы широко распространились методы, использующие антитела, меченные частицами тяжелых металлов, например коллоидного золота, железа и др.
Указанные способы позволили получить весьма ценную информацию о детальном анатомическом распределении различных синапсов, В первую очередь это помогло локализовать катехоламинергические синапсы, а также синапсы, содержащие ферментные системы с уникальными метаболитами, такими как ГАМК. В отличие от катехоламинов аминокислотные нейромедиаторы – L‑глутамат, L‑аспартат, глицин – участвуют в метаболических процессах практически всех клеток, в связи с чем выбор специфических маркеров для соответсвующих синапсов затруднен. В этих случаях обнаружение самого нейромедиатора или его ферментных систем еще не позволяет судить о природе химического синапса и тогда использует косвенный подход, например измерение высвобождаемых медиаторных аминокислот из нервных окончаний при их разнообразной стимуляции.
Общая схема рапределения нейромедиаторных связей по структурам представлена на рис. 2. Как видно из схемы, пока имеется недостаточно фактов, чтобы представить полную картину химического картирования мозга. Это требует дальнейшей большой совместной работы морфологов и нейрохимиков. Однако для ряда путей, особенно функционирующих на основе биогенных аминов, эти данные уже получены. Они локализованы преимущественно в нейронах, входящих в состав полосатого тела, вентролатерального ядра, черного вещества и голубого пятна. Аксоны этих нейронов проецируются, как правило, в гипоталамус, мозжечок, передний мозг.
Полагают, что моноаминовые нейромедиаторные пути имеют отношение к проявлению эмоций, регуляции настроения, поддержанию состояния бодрствования и др. Нарушение обмена норадреналина связывают с возникновением ряда психоэмоциональных расстройств. Воздействие на указанные пути фармакологическими средствами, компенсирующими дефицит или избыток нейромедиатора, способствует в ряде случаев снижению симптоматики шизофрении, маниакально-депрессивных состояний и др.
Нейроны, содержащие дофамин, сосредоточены в областях среднего мозга, особенно их много в черной субстанции и вентролатеральной покрышке. Многие из этих нейронов посылают свои аксоны в передний мозг, где они участвуют в развитии эмоциональных реакций. Важную роль дофаминовая медиаторная система играет в регуляции сложных двигательных функций.
Обнаружено, что нарушения дофаминергических путей приводит к затруднению движений, особенно стереотипных, к возникновению непроизвольного дрожания и скованности мышц, т.е. к появлению характерных симптомов паркинсонизма. Фармакологические препараты, содержащие метаболиты дофамина, проникающие в головной мозг, способны смягчить проявление болезни. Гиперфункция дофаминергической системы связана с механизмами шизофрении.
Моноаминовый нейромедиатор серотонин сосредоточен в области ствола мозга, где находятся так называемые «ядра шва». Нейроны этого центра проецируются в гипоталамус, таламус и другие области мозга.

Как полагают, они участвуют в терморегуляции, сенсорном восприятии и процессах сна.
Лекарственные средства, которые способны частично удалить из синапсов моноаминовые нейромедиаторы, вызывают депрессию, тогда как все препараты, применяемые для лечения клинической депрессии, обычно повышают содержание этих нейромедиаторов, усиливают их действие.
Ацетилхолин широко представлен в разных отделах нервной системы, основное его количество находится в периферических нервно-мышечных синапсах, рецепторы которых относятся к категории так называемых никотиновых. В ЦНС ацетилхолин сосредоточен. преимущественно в базальных ганглиях, таламусе и сером веществе. Соответствующие рецепторы в мозге относятся главным образом к категории мускариновых. Вставочные холинергические нейроны обнаружены в хвостатом ядре, переднем роге латерального желудочка, которые являются одними из наиболее богатых ацетилхолиновых мозговых структур.
Полагают, что АХ в подкорковых структурах участвует в тонкой регуляции сложных двигательных функций, в частности в механизмах инициации движения, двигательных стереотипах и др. Поражение холинергической иннервации в структурах мозга сопровождается нарушением и извращением двигательных функций. Так, например, при паркинсонизме наряду с нарушением дофаминергической трансмиссии отмечается гиперактивность некоторых холинергических систем мозга. Поэтому для лечения этого заболевания используют препараты, содержащие холинолитики, которые снижают уровень ацетилхолина или стимулируют работу ацетилхолинэстеразы. Эти вещества вводят совместно с аналогами L‑ДОФА для компенсации дефицита дофаминергических путей. Нарушение холинергической иннервации характерно и для ряда сенильных болезней мозга.
Поражение холинергической передачи в периферической нервной системе, в частности нервно-мышечных синапсах, связано с симптомами «усталости» или «слабости» мышц. Полагают, что в основе тяжелого заболевания – миастении гравис – лежит аутоиммунный процесс. Организм вырабатывает аутоантитела, которые блокируют функцию холинорецепторов.
Наиболее широко распространеными в ткани мозга рецепторами и, соответственно, нейромедиаторами являются некоторые аминокислоты. Центральное место среди них занимает L‑глутаминовая кислота основной возбуждающий нейромедиатор. Глутаматергические синапсы распространены в коре головного мозга, гиппокампе, полосатом теле и гипоталамусе. Нисходящие глутаматергические пути обнаружены практически во всех структурах головного мозга, проекции которых идут от коры к подкорковым структурам. Выявление глутаматергических связей в головном мозге проводится преимущественно методом физиологической идентификации по. высвобождению нейромедиатора. В последние годы на основе изучения структуры и свойств глутаматных рецепторов появилась возможность визуализации нейрорецепторов глутамата с помощью моноклональных и политональных антител,
Нейрорецепторы глутамата располагаются кластерами на постсинаптической мембране. Выявление глутаматных рецепторов на мембране клеток с помощью иммуногистохимических методов является более надежным способом идентификации глутаматергических связей по сравнению с другими методами.
Нарушение глутаматергической медиации связано с целым рядом патологических состояний нервной системы: эпилепсией, расстройствами вестибулярной системы, ишемическими проявлениями и др. Глутаминовая кислота и некоторые ее аналоги используются в качестве терапевтического лекарственного средства при хронической недостаточности аминокислотного обмена, вегетососудистой дистонии и эпилепсии.
Присутствие в разных структурах мозга ГАМК – первого по значимости тормозного нейромедиатора показано методами авторадиографии. Топографическое распределение самого радиоактивно меченного нейромедиатора или образующего его фермента – глутаматдекарбоксилазы – в головном мозге неравномерно. К областям, содержащим наиболее высокую концентрацию ГАМК, относятся черное вещество, бледный шар, гипоталамус и мозжечок. Аминокислота содержится преимущественно в сером веществе головного и спинного мозга.
Данные о функциональной роли ГАМК-ергической передачи в головном и спинном мозге постоянно обогащаются новыми фактами. Она принимает участие в регуляции моторной активности, поддержании судорожного порога, формировании эмоционального поведения. ГАМК-ергическая система участвует в осуществлении условных рефлексов, организации процессов обучения и памяти у млекопитающих. При этом она тесно взаимодействует с другими медиаторными системами мозга: дофаминергической, холинергической и глутаматергической.
Имеется очень большое количество данных о вовлечении системы ГАМК в механизмы многих метаболических расстройств нервной системы. Установлено, что нарушения этой системы связаны с прявлениями эпилепсии, хореи Гентингтона, паркинсонизма и некотрых других поражений экстрапирамидной системы. При терапевтическом применении соединений, содержащих эту аминокислоту или ее аналоги, обнаруживаются позитивные клинические эффекты. Имеются данные о благоприятном влиянии производных ГАМК на больных эпилепсией, хореей Гентингтона и паркинсонизмом. Эти же препараты способны усиливать дыхание, энергетический обмен нервной ткани, улучшать показатели мозгового кровообращения и метаболизма глюкозы.
Как упоминалось, методология гистохимического исследования локализации ГАМК или образующего ее фермента – глу-таматдекарбоксилазы – неприменима для выявления других тормозных систем – глицина и таурина и их рецепторов. Анализ их распределения производят, как правило, с использованием методов электрофизиологической регистрации высвобождения нейропередатчика из нервных окончаний при их разнообразной стимуляции.
Глицин и его рецепторы локализованы в зонах моста, продолговатого мозга и серого вещества спинного мозга, включая передние и задние рога. Авторадиографически была установлена локализация участков высокоаффинного захвата глицина преимущественно в аксо-аксональных и аксодевдритных синапсах. Скопление гранул отмечены также вокруг клеточных тел спинальных мотонейронов. У больных с некоторыми врожденными метаболическими аномалиями, связанными с повышением содержания глицина в ткани мозга и крови, может развиваться гиперглицинемия, которая сопровождается симптомами нарушения некоторых психоэмоциональных функций. Полагают, что такие расстройства могут быть следствием поражения обычных путей деградации глицина в нервной клетке.
Интересные данные были получены для таурина. Уровень таурина в разных зонах мозга оказался практически одинаковым, за исключением следующих структур: медиального коленчатого тела, гипофиза и шишковидной железы. Общая концентрация таурина и цистеинсульфонатдекарбоксилазы в спинном мозге и таламических ядрах совпадает с количеством ГАМК в этих структурах, однако локализация их по зонам внутри структур существенно различается. Все исследователи таурина сходятся во мнении о его необычайно высоком содержании в коре мозжечка, которое почти в 5 раз превышает уровень ГАМК в этой структуре. Показано, что таурин локализуется преимущественно в звездчатых нейронах молекулярного слоя. Это позволило предположить существование тауринергических нейронов. Вместе с тем авторадиографическое изучение распределения таурина свидетельствует и о его преимущественно глиальной локализации.
Существует достаточно веские аргументы в пользу того, что таурин является также важным компонентом питания живых организмов, так как он не синтезируется у млекопитающих, включая человека. Клинически тауриновый дефицит может выражаться в эпилептических припадках, наследственной атаксии Фридрейха, зрительной дисфункции, называемой в просторечии «куриной слепотой», и др. Широко обсуждается возможность участия таурина в патогенезе судорожно-пароксизмальных состояний. Выяснилось, что таурин, введеный в желудочки мозга крысы, подверженной судорогам, является более мощным, чем ГАМК, противосудорожным агентом. Однако в клинической практике таурин не проявляет стабильных противосудорожных эффектов и пока не нашел широко применения.
3.7 Характеристики индивидуальных медиаторов
Ацетилхолин
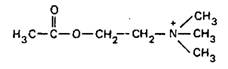
Предшественником АХ. служит холин, потребляемый с пищей. Холин поступает в холинергические нейроны с помощью специфической системы транспорта. Синтез АХ происходит в цитоплазме с участием холинацетилтрансферазы:
![]()
Затем АХ поступает в синаптические пузырьки. После экзоцитоза АХ в синаптическую щель он подвергается инактивации с участием ацетилхолинэстеразы:
![]()
АХ является преимущественно возбуждающим нейромедиатором, реже – тормозным.
У млекопитающих скопления холинергических нейронов локализуются в следующих отделах мозга: медиальное ядро перегородки, диагональная связка, базальное гигантоклеточное ядро, ядра моста. Аксоны этих нейронов проецируются на гиппокамп, проходят через кору больших полушарий. Холинергические нейроны головного мозга участвуют в таких функциях, как память,
регуляция движения, уровень бодрствования. Холинергические синапсы мозга содержат преимущественно мускариновые рецепторы. В спинном мозге АХ является нейромедиатором в синапсах, образуемых а-мотонейронами на клетках Реншоу. В вегетативной нервной системе АХ служит нейромедиаторм во всех преганглионарных нервных окончаниях симпатической и парасимпатической нервной системы – через посредство никотиновых холинорецепторов; во всех постганглионарных парасимпатических нервах, постганглионарных симпатических нервах потовых желез через посредство мускариновых холинорецепторов. АХ осуществляет через посредство никотиновых холинорецепторов функцию нейромедиатора в нервно-мышечных синапсах, образуемых соматическими эфферентными нервами в скелетных мышцах. Среди беспозвоночных АХ выявлен в качестве нейромедиатора у плоских и кольчатых червей, у моллюсков.
Моноамины. К моноаминовым медиаторам относятся катехоламины, а также серотонин и гистамин В группу катехоламинов входят норадреналин, адреналин, дофамин, ок-топамин.
Предшественником катехоламинов является L‑тирозин, который организм получает в составе пищи, а также может синтезировать в печени из фенилаланина, потребляемого с пищей. L‑тирозин поступает в нервное окончание посредством активного транспорта. Ниже приведена схема синтеза катехоламинов:

Катехоламины депонируются в оптически плотных крупных синаптических пузырьках. Деградация секретированных катехоламинов происходит с участием моноамино-оксидаз и катехол-О-метилтрансферазы. Инактивация синаптического норадреналина после его экзоцитоза осуществляется также посредством обратного поглощения в нервные окончания, т.е. путем активного трансмембранного транспорта. Норадреналин

Тела норадренергических нейронов в ЦНС млекопитающих находятся в стволе мозга, главным образом в мосте мозга, в продолговатом мозге и ядре одиночного тракта. Многочисленные нейроны голубого пятна образуют диффузные проекции большой протяженности, достигающие практически всех отделов ЦНС – коры больший полушарий, лимбической системы, таламуса, гипоталамуса, спинного мозга.
Нисходящие норадренергические пути спинного мозга участвуют в регуляции мышц-сгибателей и сосудистого тонуса. В вегетативной нервной системе норадреналин является нейромедиатором постганглионарных симпатических нервов. В мозговом слое надпочечников высвобождаются норадреналин и адреналин. В ЦНС норадреналин является в ряде отделов преимущественно тормозным нейромедиатором, например в коре больших полушарий, реже – возбуждающим, например в гипоталамусе.
У беспозвоночных НА отсутствует или имеется в малых количествах. Адреналин
В головном мозге млекопитающих количество адренергических путей является гораздо более ограниченным по сравнению с норадренергическими. Тела нейронов, содержащие фе-нилзтаноламин‑1Ч-метилтранеферазу, находятся в нижних отделах моста и в продолговатом мозге. Нисходящие пути достигают центрального серого вещества и ядер гипоталамуса.
Нейромедиаторная роль адреналина сомнительна; нейромедиатором адренергических нейронов является, очевидно, НА. Адреналин высвобождается диффузно и выполняет роль модулятора.
У беспозвоночных адреналин, так же как и НА, почти отсутствует.
Дофамин
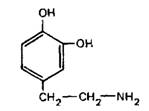
Тела дофаминергических нейронов находятся в среднем мозге, обонятельной луковице, гипоталамусе и перивентрикулярной области продолговатого мозга. Дофаминергические тракты соединяют черную субстанцию с неостриатумом, вентральную покрышку с лимбической системой и с лобной корой, аркуатное ядро гитоталамуса со срединным возвышением. Дофамин служит нейромедиатором амакриновых клеток сетчатки. Дофамин выполняет нейромедиаторную функцию и у беспозвоночных.
Октопамин
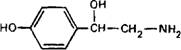
Октопамин нейромедиатор, характерный для беспозвоночных. По отношению к мозгу позвоночных он рассматривается как «ложный медиатор, не опосредующий физиологические эффекты.
Серотонин

Предшественником 5‑НТ является незаменимая аминокислота триптофан, потребляемая с пищей. Синтез 5‑НТ происходит вне секреторных гранул, которые поглощают 5‑НТ с помощью высокоаффинного переносчика:

После экзоцитоза 5‑НТ инактивируется путем активного обратного транспорта или подвергается окислительному дезаминированию с образованием 5‑гидроксииндолуксусной кислоты.
Серотонинергические нейроны составляют ядра шва в ростральной части моста мозга; эти нейроны дают проекции к лимбической системе, базальным ганглиям, коре больших полушарий. В продолговатом мозге находятся серотонинергические нейроны, аксоны которых образуют нисходящие пути в ствол мозга и в спинной мозг. Кроме того, серотонин обнаружен в нейронах гипоталамуса, черной субстанции, спинном мозге. Серотонин широко распространен и у беспозвоночных.
5‑НТ играет важную роль в регуляции эмоционального поведения, двигательной активности, пищевого поведения, сна, терморегуляции, участвует в контроле нейроэндокринных систем. 5‑НТ может выполнять не только роль нейромедиатора, но и роль нейромодулятора.
Гистамин

Гистамин образуется с помощью фермента гиствдиндекарбоксил азы

Деградация гистамина происходит с помощью моноамино-оксидазы или гистаминазы.
Гистаминергическая система весьма своеобразна по своей локализации и функциям. Нейроны, продуцирующие гистамин, сосредоточены в очень ограниченной области мозга туберо-мамиллярных ядрах заднего гипоталамуса. Эта группа нейронов посылает свои эфферентные волокна практически во все отделы мозга. Везде обнаруживаются как постсинаптические, так и пресинаптические рецепторы гистамина. Важным условием исследования локализации элементов гистаминергической системы явилось открытие специфического ингибитора синтеза гистамина – а-флюоро-метил-гистамина. Столь широкая локализация рецепторов гистамина позволяет понять чрезвычайно разнообразные функции этой системы. Гистамин уменьшает продолжительность ортодоксальной фазы сна и облегчает пробуждение. Он стимулирует двигательную активность, половое поведение и в то же время подавляет восприятие боли. Усиливая жажду, он в то же время подавляет пищедобывательное поведение. Гистамин входит в число факторов, которые через центральные механизмы участвуют в повышении давления крови, в терморегуляции и в управлении энергетикой мозга стимуляция гидролиза гликогена. Интересно, что гистамин реализует эти функции не только в рамках классических синапсов, часть гистамина выделяется так называемыми открытыми нервными окончаниями и способна к распространению по межклеточным жидкостям, в том числе через ликвор.
Аминокислоты. Аминокислотные нейромедиаторы в соответствии с их функцией делятся на две группы – возбуждающие аминокислоты и тормозные.
Глутамин поступает в организм с пищей. В нейроны он поступает из глии и служит предшественником для синтеза глута-мата, аспартата и ГАМК:

С пищей в организм поступает также аспартат, глицин, таурин.
Глутаминовая кислота

Глутамат обнаруживается во всех отделах ЦНС, очевидно, благодаря тому, что он является не только нейромедиатором, но и предшественником других аминокислот. Тела глутаматер-гических нейронов находятся в коре больших полушарий, обонятельной луковице, гиппокампе, черной субстанции, мозжечке, сетчатке. Глутаматергические синапсы существуют в миндалине, стриатуме, на клетках-зернах мозжечка. В спинном мозге глутамат сосредоточен в первичных афферентных волокнах дорсальных корешков.
Глутамат возбуждающий нейромедиатор в мозге животных, а также в нервно-мышечных синапсах ракообразных и насекомых.
Аспаоагиновая кислота

Наиболее высокое содержание аспартата найдено в среднем мозге. В спинном мозге аспартат содержится в дорсальном и вентральном сером веществе. Предполагается нейромедиатор-ная роль аспартата в возбуждающих интернейронах, которые регулируют различные спинномозговые рефлексы.

Широко распространена в ЦНС млекопитающих – она выявляется примерно в 50% всех нервных окончаний мозга. ГАМК представляет собой основной тормозной нейромедиатор в ЦНС. В коре больших полушарий имеется большое тсоличество ГАМК-ергических тормозных интернейронов. ГАМК находится в нейронах стриатума, дающих проекции на черную субстанцию, в нейронах мозжечка. В желатинозной субстанции задних рогов спинного мозга присутствуют ГАМК-ергические аксо-аксонные синапсы на первичных афферентных волокнах; эти синапсы опосредуют деполяризацию и ослабление секреции нейромедиатора – пресинаптическое торможение. Таким образом осуществляется тормозная регуляция ot‑мотонейронов. Высокие концентрации ГАМК найдены в горизонтальных клетках сетчатки; предполагается, что ГАМК обеспечивает обратную связь и латеральное торможение в слое горизонтальных клеток.
Глицин
HOOC-CH2-NH2
Эта аминокислота выполняет нейромедиаторную роль прежде всего в спинном мозге млекопитающих, где она опосредует постсинаптическое торможение мотонейронов, высвобождаясь из окончаний клеток Реншоу. Глицин является нейромедиато-ром также в тормозных интернейронах промежуточного мозга и ретикулярной формации продолговатого мозга. Наряду с ГАМК глицин прослеживается в сетчатке.
Таурин
HS03-CH2-CH2NH2
В качестве гипотетического тормозного нейромедиатора у млекопитающих следует назвать таурин, который содержится в головном и спинном мозге. В сетчатке таурин, возможно, служит нейромедиатором * тормозных синапсах внутреннего плексиформного слоя.
Пурины. В последнее десятилетие установлено, что нейромедиаторами служат и разнообразные пурины. Существуют два главных типа пуринергической трансмиссии. В первом основным нейромедиатором служит аденозин, во втором – АТФ и более сложный его дериват – диаденозинтетрафосфат
Аденозиновая трансмиссия включает медленные, метаботроп-ные рецепторы, модулирующие синтез цАМФ. АТФ-трансмис-сия осуществляется частично через быстрые, канальные рецепторы, модулирующие ионные потоки, особенно Са+.
Физиологические эффекты аденозина изучены довольно основательно. Они включают сопряжена со стимуляцией сократимости сердечной мышцы и опять-таки гипотензивным действием.
Пептидные медиаторы. Нейропегттиды составляют весьма многочисленную и полифункциональную группу. Некоторые нейропептиды удовлетворяют критериям нейромедиаторов. С известной долей осторожности сюда можно отнести вещество Р, вазоактивный интестинальный полипептид, сомато-статин, нейропептид Y, люлиберин. Гораздо более значительное число нейропептидов обладает свойствами нейромодуляторов.
Вещество Р. Оно оказалось первым веществом пептидной природы, у которого были обнаружены свойства нейромедиатора. Вещество Р содержится в телах первичных сенсорных нейронов спинномозговых ганглиев, депонируется в оптически плотных пузырьках, перемещается к пресинаптическому окончанию посредством быстрого аксонного транспорта, высвобождается из сенсорных нейронов под влиянием деполяризации при условии наличия Са* в среде. При аппликации на дорсальные рога спинного мозга вещество Р вызывает мощное возбуждение сенсорных нейронов второго порядка. После введения животным нейротоксина капсаицина, который обусловливает избирательную дегенерацию немиелинизированных первичных афферентов, происходило снижение содержания вещества Р в дорсальной части спинного мозга и'исчезновение медленных потенциалов дорсальных корешков.
■ Таким образом, вещество Р можно считать нейромедиато-ром пресинаптических окончаний С-волокон первичных сенсорных нейронов, образующих синапсы на сенсорных нейронах второго порядка в задних рогах. Этот – процесс участвует в восприятии болевых сигналов.
Как известно, первичные сенсорные нейроны образуют кроме центральных синапсов периферические синапсы в гладких мышцах дыхательных путей, кровеносных сосудов, желудочно-кишечного тракта, органов мочеполовой системы. Вещество Р высвобождается и в этих синапсах, инициируя медленные возбуждающие постсинаптические потенциалы, которые связаны с регуляцией тонуса гладких мышц.
Наряду с функцией возбуждающего нецромедиатора первичных сенсорных нейронов вещество Р может оказывать модулирующее влияние, в частности, усиливая десенситизацию никотиновых холинорецепторов. Наиболее характерным видом сосуществования с класическим нейромедиатором считается сочетание вещество Р + серотонин, причем вещество Р угнетает вызываемое деполяризацией высвобождение серотонина из срезов спинного мозга, а серотонин потенциирует высвобождение вещества Р. Вещество Р может сосуществовать и с другими медиаторами, классическими и пептидными: АХ, катехоламина-ми, ГАМК, ко-кальцигенином, вазоактивным интестинальным полипептидом, холецистокинином, нейротензином, соматоста-тином, опиоидными пептидами.
Вазоактивный интестинальный полипептвд Присутствие и высвобождение ВИП зарегистрировано во многих отделах нервной системы, прежде всего – в коре больших полушарий и в вегетативной нервной системе. Предполагают, что ВИП играет роль нейромедиатора в постганглионарных нейронах вегетативной нервной системы, которые участвуют в расслаблении гладких мышц кровеносных сосудов, дыхательных путей, кишечника. Электрофизиологическим отражением этих процессов являются так называемые медленные неадренергические не-холинергические тормозные постсинаптические потенциалы.
Согласно более ранней концепции Дж. Бэрнстока, нейроме-диатор неадренергических нехолинергических трансмуральных нервов кишечника имеет пуринергическую природу. При электрическом трансмуральном раздражении изолированных препаратов гладких мышц толстой кишки. и дна желудка наблюдалось высвобождение ВИП, которое коррелировало с силой расслабления мышцы. Высвобождение ВИП зарегистрировано при электрическом раздражении нервов трахеи и бронхов; обработка препарата специфическими антителами к ВИП предотвращало медленное расслабление трахеи морской свинки как в ответ на ВИП, так и в ответ на раздражение нервов. При высоких концентрациях ВИП развивалась тахифилаксия медленных ответов на раздражение нервов.
Наряду с возможной нейромедиаторной функцией достаточно определенно вырисовывается роль ВИП в качестве нейромоду-лятора, взаимодействующего с мускариновыми холинорецеп-торами в центральной и периферической нервной системе. В постганглионарных симпатических нейронах подчелюстной железы кошки обнаружена реципрокная регуляция высвобождения АХ/ВИП посредством обратной связи через мускарино-вые холинорецепторы и рецепторы ВИП. АХ ингибирует высвобождение ВИП, причем атропин блокирует этот эффект. В свою очередь, ВИП подавляет высвобождение АХ.
Хроническое введение крысам атропина сопровождалось значительным увеличением количества участков связывания – НА, вызываемое электрическим раздражением симпатических нервов семявынося-щего протока крысы; одновременно происходит подавление вызванных сокращений гладких мышц протока. Нейропептид Y усиливает тормозное влияние агониста а2-адренорецепторов кло-нидина на вызываемое деполяризацией высвобождение – НА из синаптосом продолговатого мозга. Предполагается взаимодействие между пресиналтическими а2‑адренорецепторами и рецепторами нейропептида Y, которое ведет к повышению чувствительности а2-адренорецепторов. Интересно, что клони-дин оказывает тормозное влияние на высвобождение нейропептида Y в ответ на раздражение преганглионарных симпатических нервов, тогда как антагонист а^-адренорецепторов усиливает высвобождение нейропептида YT Клонидин уменьшает связывание – нейропептида Y в срезах продолговатого мозга крыс, а нейропептид Y снижает связывание клонидина за счет увеличения константы диссоциации комплекса лиганд–рецептор без изменений количества рецепторов. Такое взаимодействие между рецепторами служит примером синаптической гете-рорегуляции.
Постсинаптическое модулирующее влияние нейропептида Y выражается в том, что он усиливает сократительные ответы гладких мышц семявыносящего протока и кровеносных сосудов на субмаксимальные концентрации НА.
Наряду с нейромодуляторной функцией не исключается и нейромедиаторная роль нейропептида У, который способен оказывать прямое сосудосуживающее влияние. После истощения катехоламинов путем введения резерпина в сочетании с перерезкой преранглионарных симпатических нервов селезенки кошки раздражение нерва селезенки сопровождается сужением сосудов. Предполагается, что этот неадренергический эффект опосредуется высвобождением нейропептида Y.
Люлибирин. Аналог гипофизарного ЛГ-РГ млекопитающих, близкий по свойствам к ЛГ-РГ костистых рыб, обнаружен иммуноцитихимическими методами в преганглионар-ных волокнах симпатических ганглиев лягушки, где он высвобождается Са+-зависимым образом в ответ на электрическое раздражение. При аппликации на симпатические нейроны ЛГ-РГ вызывает деполяризацию, идентичную позднему медленному ВПСП. Антагонисты ЛГ-РГ подавляют как деполяризаци-онные ответы на аппликацию ЛГ-РГ, так и поздние медленные ВПСП без изменений быстрых холинергических ВПСП. Таким образом, ЛГ-РГ удовлетворяет критериям нейромедиатора поздних медленных ВПСП в синапсах преганглионарных симпатических нервов.
Эта ситуация представляет собой интересный случай сосуществования двух нейромедиаторов классического и пептидного. Взаимоотношения между этими двумя нейромедиато-рами поясняет схема. В 9–10‑м паравертебральном симпатическом ганглии существуют два вида нейронов: В-клет-ки, которые снабжаются волокнами 3–5‑го спинальных нервов, и С-клетки, которые иннервируются волокнами 7–8‑го спинальных нервов. Электрическое раздражение 3–5‑го нервов вызывает холинергические быстрые ВПСП только в В-клет-ках. При раздражении 7–8‑го нервов возникают холинергические быстрые ВПСП только в С-клетках и поздние медленные ВПСП как в С-клетках, так и в В-клетках, причем в В-клетках поздние медленные ВПСП появляются примерно на 15 мс позднее, чем в С-клетках. После перерезки 3–5‑го нервов холинергические потенциалы в В-клетках исчезают, тогда как раздражение 7–8‑го нервов продолжало вызывать в В-клетках поздние медленные ВПСП. Очевидно, ЛГ-РГ высвобождается совместно с АХ из пресинаптических окончаний на С-клетке, а затем диффундирует на расстояние порядка 50 мк к В-клетке, которая не имеет синаптического контакта с С-клеткой.
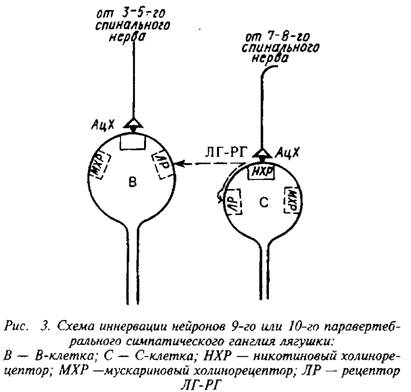
Исследование поздних медленных ВПСП в симпатическом ганглии позволило выдвинуть паракринную гипотезу действия пептидных медиаторов, которая включает два положения.
1. Нейропептиды действуют в нервной системе как паракрин-ные гормоны, т.е. могут диффундировать во внеклеточное пространство, достигая клетки-мишени, относительно удаленное от места секреции. 2. Нейропептиды, высвобождаемые из нервных окончаний, могут воздействовать и на клетки, не имеющие синаптического контакта с этими нервными окончаниями. Следовательно, классические морфологические критерии синаптического контакта непригодны для идентификации нейронов-мишеней пептидных медиаторов; наиболее важным критерием нейронов-мишеней здесь служит локализация рецепторов ней-ропептида. При этом из нервных окончаний могут высвобождаться несколько медиаторов, каждый из которых имеет свой «химический» адрес.
Нитроксид. В течение последних пяти лет накапливаются данные о возможной роли N0 в межклеточной передаче сигнала. Начало этому направлению было положено выявлением в тканях животных биохимических систем, способных генерировать N0, используя в качестве исходного соединения аргинин, а также идентификация N0 как одного из главных факторов релаксации сосудов.
Далее было показано образование N0 в ткани мозга, выявлен ряд проявлений нейрологической активности N0 и, наконец, установлен механизм его действия – посредством активации гуаншгатциклазы. Классическая схема – образование и/или накопление нейромедиатора в термикали, его выход в синапти-ческую щель после поступления импульса и включения рецептора – не подходит для описания процессинга и эффектов NO. Первый, наиболее изученный вариант, состоит в том, что при интенсивной импульсации глутаматергических синапсов синтез N0 интенсифицируется в постсинаптической зоне. Оттуда N0 выходит в межнейрональную жидкость и может активировать гуанилатциклазу, повышая уровень цГМФ в терминалах и в глии. Результаты этих процессов могут быть различны в зависимости от места первичного образования N0. В их число входит участие N0 в феноменах пластичности нейронов, консолидации памяти и т.п.
Второй вариант схемы допускает синтез N0 в терминалях и, далее, воздействие на гуанилатциклазу в постсинаптических зонах гладкой мускулатуры сосудов, тонкой кишки и некоторых других образований. Ряд исследователей полагают, что роль NO в мозге состоит главным образом в релаксации сосудов и усилении кровоснабжения в тех именно случаях, когда особенно интенсивно функционирует глутаматергическая система. Иначе говоря, ведущей, с этой точки зрения, предполагается трофическая функция.
Интересно, наконец, отметить данные о том, что малые концентрации N0 оказывают в мозге нейропротекторное действие, а относительно большие участвуют в повреждении нейронов.
Предстоит, очевидно, еще большой цикл исследований для уточнения функций и механизмов действия N0.
Выводы
1. Большинство синапсов в нервной системе млекопитающих является химическими.
2. Процесс передачи сигнала в химическом синапсе осуществляется посредством освобождения нейромедиаторов из пресинаптических нервных окончаний.
3. К нейромедиаторам относятся в настоящее время 4 группы веществ: моноамины, аминокислоты, пуриновые нуклеоти-ды, пептиды.
4. В индивидуальном нейроне синтезируется, как правило, несколько нейромедиаторов различной химической природы.
5. Существует 2 типа механизмов преобразования химического сигнала в синапсе: ионотропный и метаботропный.
6. Кроме нейромедиаторов существует обширный класс соединений – нейромодуляторов, регулирующих уровень синаптической передачи.
© 2010 Интернет База Рефератов