
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Реферат: Нейроны как проводники электричества. Физиология синапсов
Реферат: Нейроны как проводники электричества. Физиология синапсов
БЕЛОРУССКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
кафедра физиологии человека и животных
РЕФЕРАТ
На тему:
«Нейроны как проводники электричества. Физиология синапсов»
МИНСК, 2008
Нейроны как проводники электричества.
По нервам (отдельным нервным волокнам) сигналы распространяются в виде потенциалов действия и электротонических потенциалов, но на разные предельные расстояния. Способность аксонов и дендритов, а также мембран мышечных клеток проводить электрические сигналы характеризуется их кабельными свойствами.
Кабельные свойства нервных проводников очень существенны для распространения сигналов в нервной системе. Они обусловливают генерацию потенциалов действия в сенсорных нервных окончаниях, или рецепторах, при действии раздражителей, проведение сигналов по аксонам, суммацию сигналов мембраной сомы нейрона.
В основу современных теорий кабельного проведения возбуждения положена гипотеза Германна о существовании круговых токов (токов Германна), текущих от невозбужденных участков мембраны к возбужденным при распространении импульса по аксону.
Из цитологии известно, что каждая возбудимая клетка ограничена плазматической мембраной, к которой примыкают окружающие клетку оболочки. Наиболее часто нервные волокна окружены миелиновой оболочкой, сформированной глиальной клеткой в ЦНС либо оболочкой, образованной Шванновской клеткой на периферии. В местах разветвления аксона, либо в начальной или конечной его части миелиновая оболочка истончается. Сама мембрана состоит из липидов и белков. Все это обусловливает высокое электрическое сопротивление мембраны клеток и высокую распределенную электрическую ее емкость. Эти характеристики определяют проводящие свойства нервного волокна.
Основные закономерности распространения потенциалов, электротонических прежде всего, по нервным волокнам получены в экспериментах на крупных аксонах кальмаров. Было обнаружено, что при нанесении прямоугольного стимула в определенной точке волокна по мере удаления от места стимуляции сигнал регистрируется с искажением. С одной стороны, происходит изменение формы его переднего и заднего фронта (запаздывание достижения максимального значения) и уменьшение его амплитуды. Первая из этих величин определяется постоянной времени, вторая – постоянной длины. Из радиофизики известно, что постоянная времени электрической цепи имеющей емкость (С) и сопротивление R, определяется формулой
τ=RC
и измеряется в секундах.
Из чего складывается сопротивление клеточной мембраны? В клетке существует три пути, по которым может течь ток в продольном направлении по аксону
а) аксоплазма
б)внеклеточная жидкость
в) сама мембрана
Внеклеточная жидкость –электролит, ее сопротивление мало. Удельное сопротивление мембраны толщиной 100 ангстрем приближается к 1000-5000 ом×см, очень велико. Удельное сопротивление аксоплазмы невелико, 200 ом×см. Емкость мембран возбудимых клеток С близка к 1 мкф/см2, но не бывает больше 7 мкф/см2. Таким образом, τ может быть 0,1-7 мс. Постоянная времени определяет скорость запаздывания развития потенциала до максимального значения и скорость запаздывания его затухания до фоновой величины.
Градиент нарастания потенциала (заряд мембранного конденсатора) определяется экспоненциальным законом:
V/V0=(1-e–t/ τ)
Величина потенциала Vt в момент времени t меньше первоначального потенциала V0 на величину, определяемую выражением (1-e–t/ τ).
Примем t=τ, тогда
Vt/V0=(1-e–1)= 1-1/e=1-1/2.7=0.63
Или 63% от первоначального.
Разряд конденсатора мембраны тоже описывается экспоненциальной формулой:
Vt/V0=e–t/ τ
Примем t=τ, тогда Vt/V0=e–1=1/2,7=0,37 или 37% от максимального спустя время t.
Если через мембрану клетки течет емкостной ток электротонического происхождения, за каждый отрезок времени, равный τ, постоянной времени, сигнал электротона увеличивается на 63 % от предыдущего при возрастании сигнала, или уменьшается до 37% от предыдущей величины при его спаде.
Ионный механизм этого явления может упрощенно быть описан таким образом. При введении в клетку положительных зарядов (деполяризация) ионы К+ начинают перемещаться по направлению к мембране, которая обладает емкостью, позволяющей эти заряды накапливать, но имеются открытые каналы утечки, пропускающие ионы и демпфирующие накопление заряда. Чтобы произошел реальный сдвиг заряда, должно пройти время. Время нужно и для восстановления первоначальной величины заряда при разряде мембранного конденсатора. Это и есть τ.
На какое расстояние может распространится электротонический потенциал по мембране нервного волокна?
Пассивное распространение сигнала электротона определяется уравнением Ux=U0×e-x/λ, в котором мы снова видим экспоненциальную зависимость.
Нетрудно произвести преобразования формулы для случая x= λ и убедится, что электротонический потенциал Ux в точке, находящейся от первоначальной на расстоянии x будет меньше первоначального U0 в e раз (до 37% от U0, т.е. ½,7), если эта точка x равна постоянной длины λ.
Постоянная длины λ, или пространственная константа поляризации зависит от сопротивления мембраны rm, сопротивления внешней среды ro и сопротивления аксоплазмы ri.
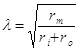
Чем больше сопротивление мембраны, меньше сопротивление среды, тем на большее расстояние передается электротонический потенциал. Уменьшение величины электротона в зависимости от расстояния называется декрементом.
Оказалось, что на постоянную длины влияет диаметр проводника, поскольку от него зависит сопротивление аксоплазмы. Поэтому толстые нервные волокна имеют большее расстояние λ, на которое способен расспространяться электротон.
Распространение электротона – прежде всего катэлектротона – важный физиологический феномен. В клетках, не генерирующих ПД (глия, эпителий, тонические мышечные волокна) за счет проведения электротона осуществляется функциональная взаимосвязь между клетками. В дендритном дереве нейронов, например, коры больших полушарий мозга, сигналы в виде электротона могут доходить от дендритов до сомы. В области синапса, зная рассмотренные закономерности, можно определить, на какое расстояние могут распространяться синаптические потенциалы.
Тем не менее, и расчеты, и прямые измерения дают очень небольшие расстояния, на которое способен распространяться электротон. Поэтому, если деполяризация в участке мембраны не достигает точки КУД, нет увеличения проницаемости для Na+, через потенциалзависимые каналы, происходят только пассивные изменения мембранного потенциала. Константа λ варьирует от 0,1 до 5 мм. Совершенно очевидно, что для связи в пределах ЦНС нужен другой механизм передачи сигналов. Эволюция его нашла. Этот механизм – распространение импульса.
Мерой проведения импульса по аксону является скорость. Скорость передачи потенциалов действия играет существенную роль в организации связей в нервной системе. Обычно быстропроводящие нервные волокна со скоростью проведения более 100 м/с обслуживают быстрые рефлексы, те, при осуществлении которых требуется безотлагательная реакция. Например, при неудачной постановке конечности (Вы оступились), чтобы избежать падения; в защитных рефлексах, инициированных повреждающим стимулом, и т.д. Для рефлексов быстрого реагирования требуется высокая скорость в афферентном и двигательном звене, до 120 м/с. Наоборот, некоторые процессы не требуют столь быстрого реагирования. Это относится к механизмам регуляции деятельности внутренних органов, где достаточно бывает скорости проведения около 1 м/с.
Рассмотрим события, связанные с распространением ПД по немиелинизированному нервному волокну (иногда с ошибкой говорят немиелиновому, но это бессмысленно, так как миелин не проводит ток, это оболочка!). Инициация волны возбуждения может быть обусловленной либо активностью рецептора (генераторный потенциал), либо синаптическими процессами. Можно вызвать ПД и электростимуляцией аксона. Если локальное смещение мембранного потенциала (деполяризация) превышает порог, достигает точки КУД, активирует потенциалзависимые натриевые каналы, ПД в виде волны деполяризации-реполяризации возникает и распространяется вдоль нервного волокна. Точка максимума ПД соответствует максимальной реверсии мембранного потенциала (овершут). Получается ситуация, при которой ПД, распространяясь по волокну, порождает перед собой токи Германна, которые разряжают мембранную емкость, приближают мембранные потенциал следующего участка мембраны к КУД, и т.д. Сзади за собой перемещающийся ПД оставляет область мембраны, находящуюся в состоянии относительной рефрактерности.
Для распространения ПД необходимо, чтобы он всякий раз порождал электротонический потенциал в соседней области, той, куда он распространяется, способный сместить мембранный потенциал на величину порога, то есть амплитуда потенциала действия должна превышать порог его возникновения в несколько раз. Отношение ПД/порог носит название гарантийный фактор (Uпд/Uпорог=5..7).
Скорость перемещения электротона и ПД по безмякотным волокнам мала и не превышает 1 м/с. У кальмара, за счет объединения в эмбриогенезе нескольких аксонов в один, что увеличивает общий диаметр проводника, скорость импульса в немиелинизированном волокне может быть до 25 м/с. У млекопитающих скорость повышена за счет миелинизации аксонов. Высокое удельное сопротивление миелина приводит к тому, что мембрана мякотных волокон приобретает высокое сопротивление и малую емкость. В перехватах Ранвье сосредоточены натриевые потенциалзависимые каналы, в приперехватных областях калиевые, ответственные за реполяризацию. Эти особенности строение приводят к тому, что сальтаторное проведение возбуждения имеет высокую надежность и высокую скорость, которые сочетаются с экономичностью (мякотные аксоны для перемещения Na+ и K+ через мембрану обходятся меньшим количеством натрий-калиевой АТФазы). Отличительное биофизическое свойство сальтаторного проведения ПД состоит и в том, что токи замыкаются через межклеточную среду, имеющую низкое сопротивление, при этом токи следуют и вдоль, и поперек волокна.
Скорость передачи импульсов по мякотному волокну зависит от диаметра последнего простым соотношением
V=К×d, где d –диаметр, а к-константа.
Для амфибий к=2, для млекопитающих к=6.
Длина участка волокна, вовлеченного в процесс передачи одного ПД равна L=t×V, где t-длительность импульса. Этот показатель важен в методическом отношении, поскольку от длины возбужденного участка нерва зависит подбор межполюсного расстояния отводящих (регистрирующих) электродов.
В нервных стволах отдельные афферентные и двигательные нервные волокна расположены в компактно упакованном состоянии. Проведение по отдельным волокнам осуществляется изолированно от соседних, может распространяться в двух направлениях от места возникновения, имеет относительно постоянную скорость в любом участке аксона (кроме окончаний) и возбуждение от нескольких источников возникновения в клетке может подвергаться алгебраическому суммированию. Диапазон различий в скоростях проведения в волокнах велик, что позволило провести несколько классификаций. Наиболее принятыми считаются классификации Эрлангера-Гассера (группы АαβγδВС) и, в меньшей степени, Ллойда (группы I, II, III).
Физиология синапсов.
Синаптическая передача – это инициация процесса возбуждения или торможения в возбудимой клетке, вызванная электрическим или химическим сигналом, поступившим от рецептирующей или нервной клетки, с помощью специализированного межклеточного контакта, называемого синапсом.
Известны два принципиально различных способа передачи сигналов от клетки к клетке – электрический и химический.
Синапсами называют специализированные структурно-функциональные образования, предназначенные для осуществления контактов между нервными клетками и эффекторными, используемых для передачи сигналов.
По принципу осуществления передачи синапсы бывают электрические и химические, смешанные химически-электрические, химические с прямым и непрямым способом рецепции медиатора. При прямом способе рецепции медиатора ионотропные рецепторы, расположенные на постсинаптической мембране, прямо активируются медиатором. При непрямой синаптической передаче рецепторы, относящиеся к классу метаботропных, реагируют на медиатор через адресацию последнего к вторичным внутриклеточным посредникам или G-белкам. Только после этого активируются ионные каналы.
По принадлежности осуществляющих контакт клеток различают нервно-мышечные, нейро-нейрональные, нейро-секреторные синапсы. Аксо-аксональные, аксо-соматические, аксо-дендритные, дендро-дендритные (если у клетки не выраженного аксона) являются разновидностями нейронейрональных синапсов.
По используемому типу медиатора бывают холинергические, адренергические (пептидергические, глутаматергические и т.д.) синапсы.
По характеру (или знаку действия) синапсы могут быть возбуждающими и тормозными.
Структура (ультраструктура, т.е. структура на уровне разрешения электронного микроскопа) синапсов сложна и менее всего напоминает простую модель, рассматриваемую в общем курсе физиологии. Однако в любом синапсе можно выделить пресинаптическую область, синаптическую щель и постсинаптическую область.
Строение и функции электрического синапса.
Для электрического синапса характерна очень узкая синаптическая щель, которая не превышает по ширине 10 нм, но близка скорее к 5 нм. Пресинаптическая и постсинаптическая мембрана у синапса с электрическим способом коммуникации имеют низкое удельное сопротивление. Это обстоятельство создает условия для прямого прохождения электрического тока из одной клетки в другую. В одной из разновидностей электрических синапсов, в щелевых контактах, или нексусах низкое сопротивление проходящему току обеспечивается наличием поперечных каналов – коннексонов, состоящих из белковых субъединиц. Они стереометрически точно расположены на пре- и постсинаптической мембране, их канальная часть имеет диаметр около 10 ангстрем. Ионы-носители тока по каналам способны проникать из одной клетки в другую. Волна деполяризации, следующая перед ПД, может вызывать смещение потенциала мембраны до КУД и инициировать появление ПД в постсинаптической клетке. Через нексусы способны проникать и некоторые низкомолекулярные соединения.
Транзит возбуждения через электрический синапс может быть с ослаблением амплитуды сигнала, поэтому введено понятие коэффициент передачи:
К.П.=Uпост./Uпрес.
Реально коэффициент передачи КП=½ означает, что амплитуда ПД ослабляется при прохождении синапса в 2 раза. Иногда синапс может проводить сигнал в двух направлениях, но коэффициент передачи в прямом и обратном направлении неодинаков. Это создает условия для того, чтобы пропускать почти без потерь деполяризацию, но практически задерживать гиперполяризующий потенциал. Говорят, что такие синапсы обладают выпрямляющими свойствами.
Иногда электрический синапс проводит только электротонический сигнал, далекий по амплитуде от порогового для достижения КУД. Но этого оказывается достаточно для процесса интеграции сигналов постсинаптической клеткой. Для электрического синапса характерна очень малая синаптическая задержка, равная сотым долям мс.
Как правило, электрические синапсы образуются между функционально однородными клетками и служат для синхронизации их деятельности. Они найдены как у беспозвоночных, так и у позвоночных животных.
Свойства электрического синапса:
а) высокое быстродействие, высокая лабильность
б) слабость или отсутствие следовых процессов –деполяризации и гиперполяризции, слабое влияние на процессы интегации воспринимающего нейрона
в) высокая надежность передачи
Химические синапсы.
Для химического синапса характерны:
1. Синаптическая задержка, продолжительностью не менее 0,5 с;
2. Отсутствие электрического тока от пре- к постсинаптической мембране.
3. Постсинаптический потенциал как результат функционирования химического синапса. Постсинаптический потенциал (ПСП) является целью функционирования химического синапса и может быть возбуждающим (ВПСП) или тормозным (ТПСП). Термины ВПСП и ТПСП применяют чаще к синапсам, образованным нейронами на нейронах. В нервно-мышечном синапсе целью синаптической передачи является формирование потенциала действия, сопряженного с последующим мышечным сокращением.
4. Увеличение проводимости постсинаптической мембраны при реализации функций синапса (ПСП в виде ТПСП или ВПСП обусловлены перемещением ионов через ионные каналы в мембране).
5. Синаптические пузырьки, или везикулы, присутствующие в пресинаптических окончаниях, специфическое окрашивание характерное для постсинаптической мембраны.
6. Зависимость процесса высвобождения, или релизинга медиатора, от входа в пресинаптическое окончание ионов Са++.
Возбуждающие химические синапсы.
Для химического синапса характерны пресинаптическая область, синаптическая щель и постсинаптическая область.
Синаптическая щель имеет у химических синапсов просвет от 20 до 50 нм. В пресинаптической области всегда имеются везикулы, содержащие медиатор (трансмиттер, нейротрансмиттер, нейромедиатор).
В рассматриваемом типе синапса из-за высокого сопротивления синаптических мембран и широкой синаптической щели электротонический потенциал и ПД не способны перейти к постсинаптической области, используя кабельные свойства мембраны. Коэффициент передачи в этом случае меньше тысячных долей, а внеклеточный шунт имеет низкое сопротивление и «уводит» заряд. По пресинаптическому волокну механизмом аксонного транспорта (400 мм/сут) осуществляется передвижение ферментных систем и предшественников для синтеза медиаторов и везикул. В синаптическом окончании всегда имеется некоторый запас готового для секреции медиатора, упакованного в везикулы.
Синтез медиаторов осуществляется с помощью ферментов, например, ацетилхолин АХ синтезируется холинацетилтрансферазой, которая переносит ацетильную группу от ацетилкоэнзима А на холин. Примерно 85% готового медиатора хранится в везикулах. Процесс синтеза и распада АХ происходит постоянно.
Выход медиатора из окончания также происходит непрерывно, это так называемый неквантовый релизинг, его интенсивность может превышать действенный, квантовый в десятки раз, но электрогенных последствий он не имеет (оказывает трофическое действие на объект иннервации), и АХ разрушается без изменения проницаемости постсинаптической мемебраны.
Квантовый выход АХ имеет электрически значимые последствия. Инициация квантового релизинга задается приходом по аксону потенциала действия, который в потерявшем миелин пресинаптическом окончании деполяризует его мембрану, что приводит к открытию потенциалчувствительных Са++ каналов. Из-за высокого электрохимического и концентрационного градиента ионы Са++ входят в пресинаптическое окончание. Кальций необходим для того, чтобы везикулы с медиатором могли соединиться с внешней мембраной и выпустить порцию (квант) медиатора в синаптическую щель путем экзоцитоза. Одновременно в синапсе может опорожниться до сотен везикул. В кванте бывает от 102 до 105 молекул АХ.
Мишенью АХ в холинергическом синапсе является комплексная белковая молекула холинорецептор. Холинорецепторы, чувствительные к никотину, относят к типу Н-холинорецепторов, к мускарину – М-холинорецепторов (метаботропных). Н-холинорецепторы расположены (экспрессированы) на мембранах мышечных волокон скелетных мышц, нейронов ЦНС и симпатических ганглиев.
Н-холинорецептор, ионотропный, состоит из 5 (иногда 7) белковых субъединиц, одна из которых дублируется (αβαγδ). Общий размер (11×8,5 нм) молекулы вдвое больше толщины мембраны. Аминокислотная последовательность белков всех субъединиц установлена, она оказалась видоспецифичной, хотя различия у близких видов животных незначительны. Продублированные α–субъединицы обладают чувствительностью к лиганду. Холинорецептор можно рассматривать в качестве ионного канала, поскольку как интегральный мембранный белок он пронизывает клеточную мембрану и имеет центральную пору. Известны 2 состояния молекулы холинорецептора – закрытое и открытое. В открытом состоянии центральная пора холинорецепторы имеет размер около 0,7 нм, что достаточно для проникновения через нее одновалентных катионов, преимущественно Na+ и К+.
После связывания АХ с Н-холинорецептором и открытия поры через постсинаптическую мембрану течет ионный ток, обусловленный перемещением ионов Na+ и К+ по электрохимическим и концентрационным градиентам. Поскольку градиент для натрия направлен внутрь клетки, а для калия - наружу, при встречном их перемещении суммарный ток оказывается способным локально сместить мембранный потенциал до КУД в нервно-мышечном синапсе или вызвать значительную деполяризацию мембраны нейрона в нейро-нейрональном синапсе. Локальный ответ в виде деполяризации в данном случае носит название ПСП – постсинаптический потенциал, или ВПСП, возбуждающий постсинаптический потенциал. Прежде часто применялось название потенциал концевой пластинки (ПКП), для нервно-мышечного синапса.
Локальный ответ в виде ВПСП подчиняется законам проведения потенциалов по мембране и может быть распространен на небольшое расстояние из-за ограничений, накладываемых емкостными и резистивными свойствами мембраны – постоянной времени и постоянной длины. Поскольку на мембране нейрона или мышечного волокна имеется множество синапсов, ответ клетки всегда складывается из активности отдельных синаптических входов.
Суммация ПКП приводит к состоянию, когда мембранный потенциал смещается деполяризацией до КУД, происходит генерация ПД. В клетку по потенциалзависимым кальциевым каналам входит кальций, он участвует в механизме мышечного сокращения.
После того, как АХ выполнил роль сигнальной молекулы и запустил конформацию холинорецептора из закрытого в открытое состояние, необходимо подготовить систему к приему следующего сигнала. Поэтому постсинаптическая мемебрана располагает механизмом инактивации медиатора. В холинергичесом синапсе инактивация АХ достигается его энзиматическим расщеплением с помощью ацетилхолинэстеразы. В других типах синапсов инактивация проходит по-другому, например, норадреналин в адренергическом синапсе подвергается обратному поступлению (захвату) в пресинаптическое окончание.
Ацетилхолинэстераза может быть заблокирована, в таком случае каналы холинорецептора постоянно находятся в открытом состоянии и управление мышцами нарушается. Инсектицидные препараты типа «Прима», «Диклофос» имеют такой принцип действия, поэтому опасны не только для бытовых насекомых-вредителей, но и для теплокровных животных.
Этапы функционирования химической синаптической передачи.
1.Синтез, хранение и транспорт медиатора в везикулах.
2.Секреция медиатора при деполяризации пресинаптической мембраны и входе ионов кальция в окончание.
3.Реакция постсинаптитческой мемебраны в виде связывания медиатора рецептором и изменении проницаемости постсинаптической мембраны для катионов.
4.Генерация постсинаптических потенциалов.
5.Инактивация медиатора.
Возбуждающие химические синапсы, образованные на нейронах, весьма многочисленны, перемежаются с тормозными, никогда не обеспечивают по одиночке достижение мембраной КУД. Нейрон способен интегрировать синаптические сигналы и выдавать на выходе, в наиболее возбудимой части клетки, например, если это мотонейрон, в аксонном холмике, ПД после проведенного анализа поступивших по синаптическим входам ПСП.
В нейро-нейрональных синапсах не только АХ может быть медиатором, чаще всего возбуждающие аминокислоты глутамат и аспартат, норадреналин, нейропептиды, АТФ и NO выполняют функции медиаторов.
Глутаматная возбуждающая синаптическая нейропередача наиболее распространена в ЦНС. Рецепция глутамата в синапсах осуществляется NMDA и AMPA (ионотропными) рецепторами, синаптические механизмы в них очень сложны и до конца не раскрыты.
Из-за того, что процессы выделения и разрушения медиатора в синапсах имеют длительное время реализции, существует синаптическая задержка при функционировании нейронных сетей. Поэтому говорят, что химический синапс работает как частотный фильтр и обладает низкой лабильностью.
Поскольку сигналы от отдельных синапсов могут суммироваться и определять суммарный заряд мембраны, возможны явления тетанического синаптического облегчения и депрессии.
Свойства химического синапса.
1.Медленная скорость передачи сигнала, большая синаптическая задержка.
2.Одностороннее проведение сигнала от пре- к постсинаптической мембране, но не наоборот.
3.Высокая надежность передачи при нормальных условиях функционирования.
4.Существование следовых процессов (следовой деполяризации и гиперполяризации, что увеличивает возможности интегрирования сигналов нейроном).
Синапсы тормозного действия.
Синаптическим торможением называется такое влияние пресинаптической нервной клетки на постсинаптическую которое сопровождается устранением или предотвращением процесса возбуждения. Тормозных синапсов на нейронах ЦНС больше, чем возбуждающих.
Тормозные синапсы могут быть электрическими. По таким межклеточным контактам возможна передача электротонического сигнала, вызывающего гиперполяризацию постсинаптической мемебраны.
Структура химического синапса тормозного типа в общем плане соответствует таковой возбуждающего, на электронномикроскопическом снимке отличить их по видимым структурным особенностям трудно. Основными медиаторами тормозных синапсов являются ГАМК, глицин, но могут использоваться АХ и другие, встречающиеся и в «возбуждающих» синапсах.
ГАМК, гамма-аминомасляная кислота, наиболее характерна для тормозных синапсов головного мозга, в спинном мозге чаще используется глицин.
После синтеза ГАМК содержится в везикулах, для высвобождения ее из пресинаптического окончания необходим Cа++. Рецепторы, чувствительные к ГАМК, связаны с открытием не катионных, а анионных каналов, проводящих ионы Cl-. Если хлор поступает в клетку по хлорным каналам, общая электроотрицательность цитоплазмы возрастает, что означает гиперполяризационное «отодвигание» мембранного потенциала от КУД. Кратковременный локальный ответ в виде гиперполяризации обозначается ТПСП тормозной постснаптический потенциал.
Таким образом, ТПСП «уводит» мембранный потенциал от КУД, интерферирует с ВПСП, блокируя («шунтируя») его действие на мембрану клетки. В генерации одного ТПСП участвует 100-200 квантов медиатора.
Рассмотренный случай является примером постсинаптического торможения.
В организме при рефлекторной деятельности бывает весьма выгодно реализовать пресинаптическое торможение. Проще предотвратить возбуждение нейрона, чем потом тормозить уже начавшееся развитие возбуждения. Такой механизм реализован в спинальном торможении болевых сигналов, и он связан с деполяризацией первичных афферентов. Деполяризация приводит к уменьшению входа кальция в пресинаптическое окончание, что сопровождается уменьшением порции выделяющегося медиатора.
Общая характеристика медиаторов.
Медиаторы, идентифицированные в настоящее время, чаще всего в химическом отношении являются аминокислотами или их производными, нейропептидами с малой молекулярной массой, или даже простыми соединениями, например, NO и CO –монооксиды азота и углерода. Как правило, это полярные молекулы. АХ был исторически первым медиатором, доказанным в качестве такового для нервно-мышечного синапса. Было время, когда энтузиасты отрицали даже возможность существования других медиаторов (милостиво разрешив норадреналину тоже быть медиатором), в том числе для синапсов на нейронах ЦНС. Сейчас, в современных представлениях, значение холинергической передачи не выше, чем остальных, поскольку в организме не может быть приоритетов ни для какой формы активности, тем более для молекулы. Наиболее распространенным медиатором, как оказалось, является глутамат. Декарбоксилированный глутамат (от молекулы отнимается один фрагмент CO2) – это уже ГАМК, наиболее распространенный медиатор тормозных синапсов. Природа весьма экономна и целесообразна!
Репертуар медиаторов в ЦНС широк и разнообразен.
Нет только «возбуждающих» и только «тормозных» медиаторов! Есть возбуждающие и тормозящие синапсы! АХ в передаче парасимпатических влияний на сердце выступает как тормозящий! АХ при стимуляции моторики кишечника вагусом возбуждает! Для NMDA рецептора ЦНС характерны сайты связывания глутамата и глицина, только обе эти аминокислоты обеспечивают функционирование синапса! Но глицин используется в спинном мозге как тормозной медиатор…
ЛИТЕРАТУРА
1. Жуков В.В., Пономарева Е.В. Физиология нервной системы: Учебное пособие / Калинингр. ун-т. - Калининград, 1999.
2. Дж.Г. Николлс, А.Р. Мартин, Б.Дж. Валлас, П.А. Фукс От нейрона к мозгу М: ЭКСМОС 2003
© 2010 Интернет База Рефератов