
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Статья: Получение мембранного белка бактериородопсина, меченного дейтерием по остаткам ароматических аминокислот L-фенилаланина, L-тирозина и L-триптофана
Статья: Получение мембранного белка бактериородопсина, меченного дейтерием по остаткам ароматических аминокислот L-фенилаланина, L-тирозина и L-триптофана
ПОЛУЧЕНИЕ МЕМБРАННОГО БЕЛКА БАКТЕРИОРОДОПСИНА, МЕЧЕННОГО ДЕЙТЕРИЕМ ПО ОСТАТКАМ АРОМАТИЧЕСКИХ АМИНОКИСЛОТ L-ФЕНИЛАЛАНИНА, L-ТИРОЗИНА И L-ТРИПТОФАНА.
@2006 г. О.В. МОСИН
Московская государственная академия тонкой химической технологии им. М.В. Ломоносова, 117571, Москва, проспект Вернадского, д. 86.
Биосинтетически получены препараты мембранного белка бактериородопсина, в которых природные ароматические аминокислоты L-фенилаланин, L-тирозин и L-триптофан селективно замещены на их дейтерий-меченные аналоги. Это достигается путем культивирования штамма галофильных бактерий Halobacterium halobium ЕТ 1001 на синтетической среде, содержащей вместо природных аминокислот химически синтезированные L-[2,3,4,5,6-2H5]-фенилаланин, L-[3,5-2H2]-тирозин и L-[2,4,5,6,7-2H5]-триптофан. Представлены данные по культивированию H. halobium ЕТ 1001 на средах, содержащих дейтерий-меченные аналоги ароматических аминокислот и выделению дейтерий-меченного бактериородопсина. Анализ степени дейтерированности аминокислот гидролизатов бактериородопсина проводили методом масс-спектрометрии электронного удара после препаративного разделения их методом обращенно-фазовой высокоэффективной жидкостной хроматографии (ВЭЖХ) в виде метиловых эфиров дансил-аминокислот и бензилоксикарбонильных производных аминокислот. Показано, что в масс-спектрах производных аминокислот гидролизатов бактериородопсина присутствуют молекулярные ионы, которые соответствуют ароматическим дейтерий-меченным аминокислотам и практически отсутствуют их нативные немеченные аналоги. Полученные данные свидетельствуют о высокой эффективности мечения бактериородопсина в этих условиях.
ВВЕДЕНИЕ.
Ретинальсодержащий белок - бактериородопсин, выполняющий функции АТФ-зависимой транслоказы в клеточной мембране галофильных бактерий Halobacterium halobium , был выделен и детально проанализирован Остерхельтом в 1970 году [1]. Несмотря на то, что бактериородопсин в настоящее время довольно хорошо изучен, он все еще остается в центре внимания исследователей по целому ряду причин. Прежде всего, благодаря своей высокой светочувствительности и разрешающей способности, он широко используется в прикладных целях как биологический фотохромный материал [2]. Кроме этого, бактериородопсин весьма привлекателен, как модельный объект для исследований, связанных с изучением функциональной активности и структурных свойств мембранных белков в составе нативных энергопреобразующих мембран.
Компьютерная модель мембранного белка бактериородопсина из H. halobium
Для получения детальной информации о структуре полипептидной цепи мембранного белка в нативной мембране целесообразно селективно вводить в белок изотопные метки, которые позволяют использовать спектральные методы высокого разрешения, такие как спектроскопия ядерного магнитного резонанса (ЯМР) [3], Раман и лазерная спектроскопия [4,5], инфракрасная (ИК) спектрометрия [6] и масс-спектрометрия (МС) [7]. В связи с этим особенно перспективны исследования с мембранными белками, селективно обогащенными стабильными изотопами, например, дейтерием по остаткам таких функционально важных аминокислот, как L-фенилаланин, L-тирозин и L-триптофан [8,9]. Это связано с тем, что указанные аминокислоты участвуют в процессе формировании хромофорного центра бактериородопсина и в его функционировании [10]. Поэтому важно получать подобные модифицированные дейтерием белки в очищенном виде и в препаративных количествах.
Целью настоящей работы было получение препаратов бактериородопсина, селективно меченных дейтерием по остаткам ароматических аминокислот - L-фенилаланина, L-тирозина и L-триптофана, а также масс-спектрометрический анализ дейтерий-меченных аминокислот в составе гидролизатов бактериородопсина после их препаративного разделения методами обращенно- фазовой высокоэффективной хроматографии (ВЭЖХ) в виде метиловых эфиров дансил-аминокислот и бензилоксикарбонильных производных аминокислот.
УСЛОВИЯ ЭКСПЕРИМЕНТА.
В работе использовали соли квалификации “х.ч”., DL-аминокислоты (Reanal, Венгрия), аденозин-и уридин-5-монофосфаты (Sigma, США.), панкреотическую телячью дезоксирибонуклеазу 1 (Fluka Chemie AG, Швейцария), додецилсульфат натрия (ДСН) (Chemapol, Чехо-Словакия). L-[2,3,4,5,6-2H5]-фенилаланин (90 ат.% 2Н), L-[3,5-2H2]-тирозин (96 ат.% 2Н) и L-[2,4,5,6,7-2H5]-триптофан (98 ат.% 2Н) (способы получения указаны в работах [ 11,12]), были предоставлены доц. кафедры биотехнологии Московской государственной академии тонкой химической технологии им. М.В. Ломоносова к.х.н. А.Б. Пшеничниковой. Для получения производных аминокислот использовали N-диметиламинонафталин-5-сульфохлорид (дансилхлорид) (Sigma, США), карбобензоксихлорид (Войковский химзавод, РФ) и диазометан. Диазометан получали из N-нитрозометилмочевины (Merck, Германия).
Бактериальные штаммы и питательные среды. Объектом исследования служил пигментированный штамм галофильных бактерий Halobacterium halobium ЕТ 1001. Штамм был получен из коллекции культур микроорганизмов Московского государственного университета им. М.В. Ломоносова. Штамм поддерживали на пептоновой среде с 2%-ным агаром [13].
Для получения немеченного бактериородопсина использовали синтетическую среду, содержащую 18 аминокислот (количества компонентов приведены в г/л): (DL-аланин 0,43, L-аргинин 0,4, DL-аспарагиновая кислота 0,45, L-цистеин 0,05, L-глутаминовая кислота 1,3, L-глицин 0,06, DL-гистидин 0,3, DL-изолейцин 0,44, L-лейцин 0,8, L-лизин 0,85, DL-метионин 0,37, DL-фенилаланин 0,26, L-пролин 0,05, DL-серин 0,61, DL-треонин 0,5, L-тирозин 0,2, DL-триптофан 0,5, DL-валин 1), нуклеотиды (аденозин-5-монофосфат 0,1, уридин-5 монофосфат 0,1), соли (NaCl 250, MgSO4 7H2O 20, KСl 2, NH4Cl 0,5, KNO3 0,1, KH2PO4 0,05, K2HPO4 0,05, цитрат натрия 0,5, MnSO4 H2O 3 10-4, CaCL2 6H2O 0,065, ZnSO4 7H2O 4 10-5, FeSO4 7H2O 5 10-4, CuSO4 5H2O 5 10-5), глицерин 1, ростовые факторы (биотин 0,1 10-3, фолиевая кислота 10 10-3, витамин В12 0,02 10-3).
В экспериментах по введению дейтериевой метки в бактериородопсин вместо L-фенилаланина, L-тирозина и L-триптофана в синтетическую среду добавляли их дейтерированные аналоги - L-[2,3,4,5,6-2H5]-фенилаланин, L-[3,5-2H2]-тирозин, и L-[2,4,5,6,7-2H5]-триптофан.
Среды стерилизовали при 0,5 ати в течении 30-40 мин, рН доводили до величины 6,5-6,7 при помощи KOH. Посевной материал выращивали в колбах Эрленмейера, объемом 250 мл (с наполнением средой до 50 мл) при 35-37 0С в условиях интенсивной аэрации и освещении лампами дневного света ЛДС-40. После суток посевной материал в количестве 5-10 % переносили в синтетическую среду, содержащую дейтерий-меченные аналоги ароматических аминокислот и культивировали в течении 4-5 суток как и при получении посевного материала.
Выделение фракции пурпурных мембран. Клетки осаждали на центрифуге Т-24 (Германия) (10000 об/мин, 10 мин.), осадок клеток суспендировали в дист. воде, экспонировали ультразвуком (3 экспозиции продолжительностью 5 мин) и вновь центрифугировали (10 000 об/мин, 10 мин). Пигменты удаляли обработкой ацетоном, липиды экстрагировали смесью хлороформ-метанол (2:1), растворитель декантировали. Полученный осадок клеток (100-150 мг) суспендировали в 100 мл буфера трис-HCL (рН 7,5), добавляли 1 мг дезоксирибонуклеазы 1 и инкубировали в течении 5-6 часов при 37 0С, затем разбавляли дист. водой до 200 мл, после чего инкубировали 15 часов при 40С. Осадок промывали водой с последующим отделением растворителя до получения бесцветных промывных вод. Контроль чистоты полученной суспензии пурпурных мембран (в Н2О) проводили на спектрофотометре “Beckman DU-6” (США) по соотношению полос поглощения при 280 нм/568 нм (e280=1,1 10 5 М-1см-1 [14] и e568=6,3 104 М-1 см-1 [15]).
Выделение бактериородопсина. Препараты пурпурных мембран (50 мг) солюбилизировали в 2 мл 0,5%-ного раствора додецилсульфата натрия (ДСН) в Н2О, выдерживали в течении 8-10 часов при 220 С, затем центрифугировали (7000 об/мин, 5 мин). Осадок отделяли, к супернатанту добавляли 5-ти кратный избыток метанола, выдерживали при 00 С в течении 12-14 часов и центрифугировали (13000 об/мин, 10 мин). Остатки ДСН удаляли, промывая осадок белка дист. водой. Выход бактериородопсина составил 20 мг.
Гидролиз бактериородопсина. Гидролиз бактериородопсина проводили двумя методами: (А)-кислотным и (В)-щелочным гидролизом. Для этого белок делили на две равные порции по 10 мг и гидролизовали в запаянных стеклянных ампулах в 30-ти кратном избытке гидролизирующего агента (24 ч, 1100 С): (А)- в 6 н. 2НCL (в 2Н2O ) с 3%-ным (по весу) фенолом и в (В) - 4 н. Ba(OH)2. После этого гидролизат, полученный в условиях (А) упаривали в роторном испарителе при 400 С. Остатки дейтеро-соляной кислоты удаляли путем выдерживания в эксикаторе над твердым NaOH. После проведения гидролиза (В), реакционную смесь суспендировали в одном объёме горячей дистиллированной воды и нейтрализовали 2 н. раствором H2SO4 до рН 7,0. Выпавший осадок сульфата бария отделяли центрифугированием (10000 об/мин, 5 мин), супернатант декантировали и упаривали в роторном испарителе при 400 С. Гидролизаты бактериородопсина, полученные в условиях (А) и (В), высушенные до постоянной массы, обрабатывали дансилхлоридом и диазометаном (или карбобензоксихлоридом).
Получение дансиламинокислот гидролизатов бактериородопсина . К 10 мг высушенных гидролизатов бактериородопсина в 2 мл 2 м. NaHCO3 (2 10-4 моль) рН 9-10 дробными порциями при перемешивании добавляли 16 мг (5,9 10-5 моль) дансилхлорида в 4 мл ацетона. Реакционную смесь выдерживали при перемешивании при 400 С в течении часа, затем подкисляли 2 н. раствором HCL до рН 3 и экстрагировали этилацетатом (3 раза по 5 мл.). Объединенный экстракт промывали дист. водой до значения рН 7,0, сушили безводным сульфатом натрия, растворитель удаляли при 10 мм. рт. ст.
Получение метиловых эфиров дансиламинокислот гидролизатов бактериородопсина. К 20 мл 40 %-ного КОН в 40 мл эфира добавляли 3 г влажной нитрозометилмочевины и перемешивали на водяной бане со льдом в течении 15-20 мин. После интенсивного газовыделения эфирный слой отделяли и промывали ледяной водой до рН 7,0, сушили безводным NaSO4 и обрабатывали им препараты дансилпроизводных аминокислот в составе гидролизатов бактериородопсина.
Бензилоксикарбонильные производные аминокислот получали реакцией Шоттен-Баумана по методике, указанной в работе [16].
Тонкослойную хроматографию (ТСХ) производных аминокислот проводили на пластинках “Silufol UV-254” (Чехо-Словакия) в системах: (A)-хлороформ-метанол-уксусная кислота (10:1:0,3) для бензилоксикарбонильных производных аминокислот и (Б)-хлороформ-метанол-ацетон (7:1:1) для метиловых эфиров дансиламинокислот. Аналитическое и препаративное разделение карбобензоксипроизводных аминокислот и метиловых эфиров дансиламинокислот в составе гидролизатов бактериородопсина проводили методом обращенно-фазовой высокоэффективной жидкостной хроматографии (ВЭЖХ) на жидкостном хроматографе “Knauer” (ФРГ), снабженным насосом “Knauer”, УФ-детектором “2563” и интегратором “С-R 3A” (Shimadzy, Япония). Использовали колонки: гиперсил ODS, 5 мкм, 3 250 мм, силасорб С18, 12 мкм, 10 250 мм. Элюирование проводили в системе растворителей по двум вариантам: (А)- вода-трифторуксусная кислота (100/0,1-0,5 об/об) и (В)- ацетонитрил-трифторуксусная кислота (100/0,1-0,5 об/об) при различных градиентных режимах, как описано в работе [17]. Бензилоксикарбонильные производные аминокислот детектировали по поглощению при 254 нм. Метиловые эфиры дансил-аминокислот детектировали по флуоресценции в УФ-свете.
Масс-спектры электронного удара производных аминокислот получены на приборе “MB-80 A (Hitachi, Япония) при энергии ионизирующих электронов 70 эВ.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ.
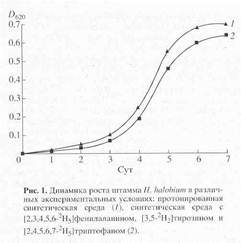
Получение дейтерий-меченного бактериородопсина. Выбор способа получения бактериородопсина был определен целью исследования, связанной с изучением принципиальной возможности получения дейтерий-меченных препаратов бактериородопсина в препаративных количествах. При выборе L-[2,3,4,5,6-2H5]-фенилаланина, L-[3,5-2H2]-тирозина и L-[2,4,5,6,7-2H5] -триптофана в качестве источников изотопных меток, авторы учитывали важность этих аминокислот в функционировании бактериородопсина, стабильность дейтерий-меченных аналогов указанных аминокислот к (1H-2H) обмену в водной среде в условиях культивирования, а также возможность их детектирования методами масс-спектрометрии. Кривые, отражающие динамику роста штамма H. halobium ЕТ 1001 на пептоновой среде (1), обычной синтетической среде(2) и синтетической среде (3), содержащей дейтерий-меченные аналоги аминокислот L-Phe, L-Tyr и L-Trp представлены на рис. 1. Как видно из рис.1, рост H. halobium ЕТ 1001 на пептоновой среде происходит лучше чем на синтетической, однако для получения дейтерий-меченного бактериородопсина пептоновая среда не подходит ввиду наличия в ней немеченных аминокислот. Поэтому пептоновая среда была заменена на синтетическую среду, в которую добавляли дейтерий-меченные аналоги ароматических аминокислот.
Способ получения дейтерий-меченного бактериородопсина представлен на схеме. Основными этапами при получении дейтерированных препаратов бактериородопсина было: культивирование штамма H.halobium ЕТ 1001 на средах, содержащих дейтерий-меченные аналоги ароматических аминокислот, разрушение клеток и получение суспензии пурпурных мембран, очистка суспензии пурпурных мембран от обломков клеток и клеточной ДНК, очистка от каротиноидов, солюбилизация белка в додецилсульфате натрия (ДСН) и осаждение конечного продукта метанолом (см. схему). При получении пурпурных мембран, клетки обрабатывали дезоксирибонуклеазой, для того чтобы разрушить клеточную ДНК. Вследствие того, что препараты пурпурных мембран содержат примеси каротиноидов и обломки клеточных стенок, было необходимо применять специальные методы очистки и выделения белка. При выделении фракции белков из бактериальных объектов необходимо учитывать наличие в них небелковых примесей. В случае богатых белками штаммов сравнительно небольшим количеством примесных соединений в них часто пренебрегают, используя в качестве белковой фракции остаток после исчерпывающего отделения пигментов и липидов экстракцией органическими растворителями. В специальных случаях для получения фракции индивидуальных белков следует прибегать к их осаждению и очистке [18]. Выделение мембранного белка заключается в его солюбилизации в растворе детергента и последующим осаждении белка. Для этого пурпурные мембраны суспендировали в растворе ДСН. Выбор ДСН в качестве детергента в данном случае оправдан, так как бактериородопсин является интегральным белком. Выделение бактериородопсина после отделения осколков клеточных стенок из раствора ДСН достигали, осаждая белок метанолом. Наиболее трудоемкой была стадия очистки бактериородопсина от каротиноидов, которая приводила к значительным потерям хромопротеида. Для этого был использован метод низкотемпературной экстракции каротиноидов ацетоном.
![]() Культивирование H. halobium ЕТ 1001на
синтетической среде, содержащей L-[2,3,4,5,6-2H5]
-фенилаланин, L-[3,5-2H2]-тирозин и L-[2,4,5,6,7-2H5]-триптофан.
Культивирование H. halobium ЕТ 1001на
синтетической среде, содержащей L-[2,3,4,5,6-2H5]
-фенилаланин, L-[3,5-2H2]-тирозин и L-[2,4,5,6,7-2H5]-триптофан.
![]()
![]()
![]() Культуральная жидкость
Культуральная жидкость
![]() Отделение биомассы
Отделение биомассы
![]() Сырая биомасса
Сырая биомасса
![]()
![]() Промывка
клеток дист. Н2О
Промывка
клеток дист. Н2О
![]()
Дезинтеграция клеток ультразвуком
![]()
![]()
![]() Ацетон. Экстракция пигментов и
липидов Экстракт пигментов и
Ацетон. Экстракция пигментов и
липидов Экстракт пигментов и
![]() Хлороформ,
липидов
Хлороформ,
липидов
метанол.
![]() Фракция суммарных белков
Фракция суммарных белков
![]() Обработка дезоксирибонуклеазой
Обработка дезоксирибонуклеазой
![]()
![]()
![]() Фракция пурпурных мембран.
Фракция пурпурных мембран.
![]()
![]()
![]()
![]() Ацетон Очистка от
каротиноидов Экстракт каротиноидов
Ацетон Очистка от
каротиноидов Экстракт каротиноидов
![]()
![]()
![]()
![]()
![]() Раствор ДСН
Осаждение белка Остатки клеточных стенок
Раствор ДСН
Осаждение белка Остатки клеточных стенок
Метанол
![]()
![]() Бактериородопсин
Бактериородопсин
![]()
![]() Гидролиз
белка
Гидролиз
белка
 Раствор 6 н. HCL с 3%-ным фенолом
Раствор 4 н. Ba(OH)2
Раствор 6 н. HCL с 3%-ным фенолом
Раствор 4 н. Ba(OH)2
1. Дансилхлорид,
![]()
![]() диазометан Модификация
аминокислот
диазометан Модификация
аминокислот
2. Карбобензоксихлорид
![]()
![]() Разделение производных аминокислот
обращенно-фазовой ВЭЖХ
Разделение производных аминокислот
обращенно-фазовой ВЭЖХ
![]() Индивидуальные производные аминокислот
Индивидуальные производные аминокислот
![]() Масс-спектрометрический анализ производных аминокислот
Масс-спектрометрический анализ производных аминокислот
Схема получения дейтерий-меченного бактериородопсина из H. halobium и определения степени включения дейтерия в аминокислоты белка.
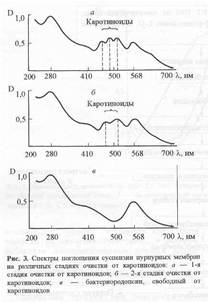
В качестве примера на рис.2,а-в изображены спектры поглощения пурпурных мембран на различных стадиях очистки от каротиноидов. Как видно из рис. 2,в, спектр поглощения бактериородопсина после экстракции от каротиноидов имеет трехполосный вид: полосы на длинах волн 568 нм и 410 нм определяются наличием хромопротеида, полоса поглощения при длине волны 280 нм определяется содержанием ароматических аминокислот в полипептидной цепи этого белка. (Для чистого бактериородопсина соотношение D280/D568 равно 2:1). Согласно предложенной схеме можно получить десятки миллиграмм дейтерий-меченного бактериородопсина, переработав 200-300 мг бактериальной биомассы. Обращенно-фазовая высокоэффективная хроматография ВЭЖХ метиловых эфиров дансилпроизводных аминокислот и бензилоксикарбонильных производных аминокислот, полученных из гидролизатов бактериородопсина показала высокие степени хроматографической чистоты выделенных аминокислот и отсутствие примесей не белковой природы в гидролизатах бактериородопсина (см. ниже).
Гидролиз дейтерий-меченного бактериородопсина. Гидролиз дейтерий-меченного бактериородопсина проводили в условиях предотвращения изотопного обмена водорода на дейтерий в ходе гидролиза и сохранения остатков триптофана в белке. Для этого были рассмотрены два альтернативных варианта проведения гидролиза- кислотный и щелочной гидролиз бактериородопсина. Кислотный гидролиз белка в стандартных условиях (6 н. HCL, 24 ч, 1100 С), как известно, приводит к полному разрушению триптофана и частичному разрушению серина, треонина и некоторых других аминокислот в белке [19], которые для наших экспериментов не играют существенной роли. Другим значительным недостатком при проведении гидролиза в HCL является изотопный обмен ароматических протонов (дейтеронов) в молекулах триптофана и тирозина [20]. Модификация метода проведения кислотного гидролиза бактериородопсина заключалась в использовании дейтерий-меченных реагентов (6 н. раствор 2НCL (в 2Н2O) ) и добавлении в реакционную смесь 3 %- ного фенола. Другой вариант гидролиза бактериородопсина заключался в использовании 4 н. раствора Ba(OH)2 (1100 C, 24 ч). В этих условиях гидролиза белка реакций изотопного обмена водорода на дейтерий в ароматических аминокислотах триптофане и тирозине не происходит, и что особенно важно - не разрушается триптофан. Оба метода гидролиза бактериородопсина показали хорошие результаты. Однако вследствие высокой гидролизующей способности гидроокиси бария и лёгкости выделения свободных аминокислот из щелочных гидролизатов путём нейтрализации гидрооксида бария серной кислотой, авторы отдали предпочтение щелочному варианту гидролиза бактериородопсина. Для последующего масс-спектрометрического анализа гидролизаты бактериородопсина обрабатывали дансилхлоридом и диазометаном (и/или карбобензоксихлоридом) с получением летучих производных аминокислот, которые затем разделяли методами обращенно-фазовой ВЭЖХ.
Масс-спектрометрический анализ метиловых эфиров дансильных производных L-[2,3,4,5,6-2H5]-фенилаланина, L-[3,5-2H2]-тирозина и L-[2,4,5,6,7-2H5]-триптофана, выделенных из гидролизатов бактериородопсина. Масс-спектр гидролизата бактериородопсина, полученный со среды, содержащей L-[2,3,4,5,6-2H5]-фенилаланин, L-[3,5-2H2]-тирозин и L-[2,4,5,6,7-2H5]-триптофан после обработки дансилхлоридом и диазометаном представлен на рис. 4. Как видно из представленных данных, в масс-спектре этого гидролизата бактериородопсина присутствуют пики молекулярных ионов обогащённых дейтерием метиловых эфиров дансилпроизводных -фенилаланина с М+. m/z 417 (вместо 412 для нативного фенилаланина), тирозина с М+. с m/z 429 (вместо 428) и триптофана с М+. m/z 456 (вместо 451). Полученные данные свидетельствует о высокой эффективности мечения бактериородопсина по остаткам дейтерированных фенилаланина и триптофана. Что касается изотопного обогащения тирозина в бактериородопсине (согласно данным масс-спектрометрии степень дейтерированности тирозина составляет 50%), авторы не исключают, что подобный результат мог быть обусловлен не вкладом биосинтеза тирозина de novo, а возможным (1Н-2Н)-обменом при выделении аминокислот или при химической модификации тирозина. Кроме вышеназванных аминокислот в масс-спектре модифицированного дансилхлоридом и диазометаном гидролизата бактериородопсина, изображенного на рис.3, четко детектируются пики молекулярных ионов производных других аминокислот, таких, как глицин (М+. с m/z 322), аланин (М+. с m/z 336), валин (М+. с m/z 364) и лейцин(изолейцин) (М+. с m/z 378). Как и следовало ожидать, эти аминокислоты в бактериородопсине не дейтерированы.
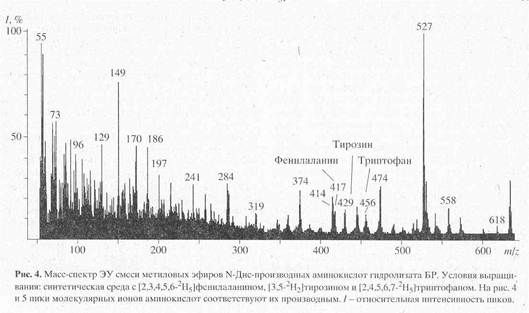
Как видно из рис.4, в масс-спектре гидролизата бактериородопсина пики молекулярных ионов, соответствующим производным ароматическим аминокислот имели недостаточно высокую интенсивность, за счет чего молекулярная область обогащения дейтерием этих соединений была сильно уширена. Поэтому было необходимо хроматографически выделять и анализировать индивидуальные дейтерий-меченные аналоги указанных аминокислот из белковых гидролизатов. Для этих целей авторы использовали метод обращенно-фазовой высокоэффективной хроматографии (ВЭЖХ), хорошо апробированный для аналитического и препаративного разделения бензилоксикарбонильных производных дейтерий-меченных аминокислот, выделяемых из других микробных объектов [17]. Этот метод был адаптирован к условиям хроматографического разделения смесей метиловых эфиров дансильных производных аминокислот бактериородопсина.
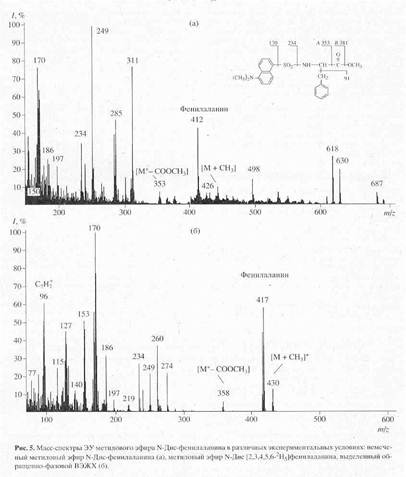
В качестве примера на рис. 5,б представлен масс-спектр чистого метилового эфира L-[2,3,4,5,6-2H5]-дансилфенилаланина, выделенного методом обращенно-фазовой ВЭЖХ из гидролизата бактериородопсина, изображенного на рис.4 (Масс спектр приведен относительно немеченного производного L-фенилаланина (а)). Доказательством наличия дейтерия в фенилаланине, хроматографически выделенном из гидролизата бактериородопсина является присутствие в масс-спектре дейтерированнного образца пика молекулярного иона дейтерированного производного L-фенилаланина (М+. с m/z 417 (вместо 412 для контрольных условий (а)) и обогащённого дейтерием бензильного фрагмента фенилаланина с m/z 96 (вместо 91 в контроле). Кроме этого, в масс спектре электронного удара метилового эфира дансил-фенилаланина зафиксированы пики фрагментов с m/z 170, 234, 250 и 353, три из которых по массе соответствуют дансильному фрагменту и продуктам его распада до N-диметиламинонафталина (эти фрагменты не дейтерированы), а низкоинтенсивный пик с m/z 353 является продуктом отщепления группы -СООСН3 из производного фенилаланина. Полученные данные свидетельствуют о том, что 5 атомов водорода в молекуле фенилаланина замещены на дейтерий, что в целом совпадает со степенью дейтерированности исходного L-[2,3,4,5,6-2H5]-дансилфенилаланина, добавляемого в среду культивирования. Согласно данным по разделению метиловых эфиров дансилпроизводных аминокислот гидролизатов бактериородопсина методом обращенно-фазовой ВЭЖХ, степень чистоты полученных метиловых эфиров L-[2,3,4,5,6-2H5]-дансилфенилаланина, L-[3,5-2H2]-тирозина и L-[2,4,5,6,7-2H5]-триптофана составляет от 96 % для производных L-фенилаланина и L-тирозина до 98% для производного L-триптофана.
Таким образом, полученные данные свидетельствуют о высокой эффективности мечения бактериородопсина по остаткам дейтерий-меченных ароматических аминокислот. В дальнейшем также планируется получать по разработанному методу полностью дейтерированные препараты бактериородопсина для реконструкции функционально активных систем мембранных белков в из очищенных компонентов в 2Н2O с участием дейтерий-меченных липидов и других дейтерированных биологически активных соединений (данные по получению полностью дейтерированного бактериородопсина будут опубликованы в отдельной статье). Эти исследования позволят дать ответ на вопрос как функционирует дейтерий-меченный бактериородопсин в составе нативных мембран в условиях, близким к адаптации организма к 2Н2О.
ЛИТЕРАТУРА.
1. Oesterhelt D. and Stoeckenius W. // Nature New Biol. - 1971. - V.233. - P.149.
2. Spudich John L. // Ann. Rev. Biophys. Chem. - 1988. - V.17. - P.193-215.
3. Creuzet F., McDermott A., Gebhard R., Hoef K., van der Spijker, Assink M. M., Herzfeld J., Lugtenburg J., Levitt M.H., Griffin R.G. // Science. - 1991. - V.251. - P.250-286.
4. Argade P.V., Kenneth J., Rothscild K.J., Kawamoto A.H., Herzfeld J., and Herlihy W.C. // Proc. Natl. Acad. Sci. USA. -1981. - V.78. - No.3. - P.1643-1646.
5. Rothschild K.J., Braiman M.S., Yi-Wu He, Marti T. and Khorana H. G. // J. of Biological Chemistry. - 1990. - P.16985-16990.
6. Otto H., Marti T., Holz M., Mogi T., Stern L.J., Engel F., Khorana H.G. & Heyn M.P. // Proc. Natl. Acad. Sci. U.S.A. - 1990. - V.87. - P.1018-1022..
7.Vetter W. , in: Biochemical Applications of mass-spectrometry (Walles G.R., and Dormor O.C.). - 1980. -First supplementary volume. - Wiley - Interscience. - N.Y. - USA. - P.439.
8. Hackett N.R., Stern L.J., Chao B.H., Kronis K.A. and Khorana H.J. // J. Biol. Chem. - 1987. - V.262. - P.9270-9277.
9. Rosenbach V., Goldberg R., Gilon C. and Ottolenghi M. // Photochem. Photobiol. - 1982. - V.36. - P.197-201.
10. Plotkin B.J., Sherman M.V. // Biochem. - 1984. - V.23. - P.5353-5360.
11. Griffiths D.V., Feeney J., Roberts G.C.and Burgen A.S. // Biochim. et Biophys. Acta. - 1976. - V.446. - P.479-585.
12.Matthews H.R., Kathleen S., Matthews K. and Stanley J. // Biochim. et Biophys. Acta. - 1977. - V.497. P.1-13.
13. Onishi H., McCance M.E., and Gibbons N.E. // Canad. J. of Microbiol. - 1965. - V.11. - P.365-373.
14. Oesterhelt D., Hess B. // Eur. J. Biochem. - 1973. - V.37. - N.1. - P.316-326.
15. Tokunada F., Ebrey T. . // Biochemistry. - 1978. - V.17. - N.10. - P.1915-1922..
16. Гринштейн Дж, Виниц М. Химия аминокислот и пептидов. - М.:Мир. - 1965. - С. 387-390. 16.
17. Егорова Т.А., Мосин О.В., Еремин С.В., Карнаухова Е.Н., Звонкова Е.Н., Швец В.И. // Биотехнология. - 1993. - Т.8. - С.21-24.
18. Oesterhelt D. & Stoeckenius W. // Methods Enzymol. - 1974. - V.31. - P.667-678.
19. Звонкова Е.Н., Зотчик Н.В., Филлипович Е.И., Митрофанова Т.К., Мягкова Г.И., Серебренникова Г.А // Химия биологически активных природных соединений. - М.: Химия, 1970. - С.65-68.
20. Cohen J.S., Putter I. // Biochim. Biophys. Acta. - 1970. - V.222. - P.515-520.
O.V. МОSIN
Moscow State Academy of Fine Chemical Technology named after M.V. Lomonosov, 117571.
PREPARATION OF BACTERIORHODOPSIN LABELED WITH DEUTERIUM ON AROMATIC RESIDUES OF AROMATIC AMINO ACIDS L-PHENYLALANINE, L-TYROSINE AND L-TRYPTOPHAN.
The samples of membrane protein bacteriorhodopsin wich contained selective deuterated analoques of aromatic amino acids such as L-phenylalanine, L-tyrosine and L-triptophan were obtained biosynthetically. It was achieved through the cultivation of a strain of halophilic bacterium H. halobium ЕТ 1001 on synthetic medium containing instead of natural amino acids the chemically synthetized L-[2,3,4,5,6-2H5]-phenylalanine, L-[3,5-2H2]-tyrosine and L-[2,4,5,6,7-2H5]-tryptophan. The data on cultivation of H. halobium on media containing the deuterated analoques of amino acids and preparation of deuterared bacteriorhodopsin are presented. The analysis of deuterium enrichment of amino acids obtained from hydrolizates of bacteriorhodopsin was perfomed by the electron impact mass-spectrometry method of methyl esters of dansyl-amino acids and benzyloxycarbonyl- amino acid derivatives after preparative separation using reverse-phase high perfomance liquid chromatofraphy (HPLC). It was shown, that in mass spectra of derivatives of amino acids obtained from hydrolizates of bacteriorhodopsin molecular ions of deuterated aromatic amino acids are presented and practically absent their native non-labelled analoques. The data obtained testified to the high efficiency of labelling of bacteriorhodopsin in those experimental conditions.
© 2010 Интернет База Рефератов