
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Учебное пособие: Предмет і завдання вивчення гістології з цитологією та ембріологією
Учебное пособие: Предмет і завдання вивчення гістології з цитологією та ембріологією
Модуль 1. Цитологія, розмноження, ембріологія
Лекція № 1. Тема: Предмет і завдання вивчення гістології з цитологією та ембріологією.
1. Гістологія вчення про тканини. Історія розвитку. Клітинна теорія.
2. Методи гістологічного дослідження.
3. Основи цитології. Історія розвитку.
4. Біологія клітин: органели, їх будова і функції.
1. Гістологія вчення о тканинах. Історія розвитку. Клітинна теорія.
Гістологія (histos — тканина, logos — вчення) — в широкому розумінні наука, яка вивча тонку і найтоншу будову, розвиток та функціонування структур організму тварини і людини.
Термін гістологія вперше введений в науку німецьким вченим К.Майєром в 1819 р. Завдання сучасної гістології полягає у тому, щоб сформувати науковий світогляд про дність органічної природи. Різноманітні методики гістологічних досліджень дають змогу вивчати організм на різних рівнях — субклітинному, клітинному, тканинному, окремих органів та цілісного організму.
Гістологія належить до морфологічних наук вчення про форму та будову організмів. Морфологія має тривалу сторію, проте лише в XVIII столітті вона оформилася у спеціальну науку. Й.В.Гете перший, у 1796 р., застосував термін «Морфологія», яку він визначив як вчення про форму, утворення і перетворення органічних тіл.
Морфологія тварин (і людини також) являє собою сукупність наук, об'єднаних спільністю об'єкту досліджень. Завданням цих наук є дослідження із застосуванням різних методів усіх проявів форм та структури. Розвиток морфології поділяють на три періоди.
1) Макроскопічний розвиток морфології припадає на античний період, середньовіччя закінчується початком XIX століття. У цей період структурні зміни в органах при різних хворобах вивчали, виходячи з особливостей їх кольору, розмірів, консистенц тощо.
2) Початок другого (мікроскопічного) періоду розвитку морфології ознаменувався створенням світлового мікроскопа (XVII—XVIII століття). Перший складний мікроскоп був виготовлений в Голландії чи Англії, тобто в найбільш передових на той час країнах. Розвитку мікроскопічного періоду сприяла розробка техніки мікроскопічного дослідження (1787—1869). Прогрес кожної науки є результатом спільних зусиль дослідників різних країн. У XVIII та на початку XIX століть проведено значну кількість досліджень, в результаті яких встановлювали мікроскопічну будову різних органів тварин і рослин.
У розвитку мікроскопічного періоду морфолог брали участь багато вчених, які надзвичайно ретельно вивчали й описували найтоншу будову різних рослинних і тваринних організмів. М.Мальпігі (1671-1675), А Левенгук (1673-1695), Сваммердам (1737), К.Ф.Вольф (1749—1769). Ц вчені поклали початок наукової мікроскопії і пробудили зацікавленість до цієї області знань, тому заслуговують великої пошани. Мікроскопічний період характеризується значними досягненнями у розвитку гістології, завдяки подальшому удосконаленню мікроскопічної техніки гістологічного дослідження. Зусиллями великої кількості вчених різних країн були виявлені найтонші детал будови клітин і тканин, основні ознаки х життєдіяльності, зроблено класифікацію. (І.Мюллер, Я.К.Пуркіньє, М.Шлєйден, Р.Ремак, А.Кьолікер, Р.Вірхов).
Великий вплив на розвиток наукової мікроскоп мала клітинна теорія Т.Шванна (1838—1839), яка містила три головних узагальнення: теорію утворення клітин, доказ клітинної будови усіх органів і частин організму й поширення цих принципів на ріст та розвиток тварин та рослин.
Створення клітинної теорії стало важливим явищем в біології, одним із вирішальних підтверджень дності усієї живої природи. Клітинна теорія значно вплинула на розвиток біології, послужила основним фундаментом для розвитку гістології, фізіології, ембріології, медицини та інших наук. Вона дала основи для матеріалістичного розуміння життя, ндивідуального розвитку, еволюційного взаємозв'язку організмів.
Основні положення клітинної теорії мають велике значення і понині.
Клітинна теорія довела, що:
· клітина є елементарною одиницею живого;
· клітини різних організмів гомологічні за своєю будовою, тобто вони мають ядро, цитоплазму, основн органели;
· розмноження клітин відбувається поділом початково клітини;
· багатоклітинні організми являють собою складні системи клітин, об'єднані в цілісні, інтегровані системи тканин та органів, підпорядкованих і пов'язаних між собою міжклітинними гуморальними й нервовими формами регуляції.
3) В 50-х роках ХХ століття інтенсивний розвиток одержала електронна мікроскопія, яка стала початком третього ультрамікроскопічного періоду розвитку морфології.
Теоретичною основою для створення електронного мікроскопа була теорія де Бройля (1924) про хвильову природу речовин та дослідження Буша (1926), який показав, що електричні та магнітні поля, що мають обертальну симетрію, діють як лінзи.
Перший електронний мікроскоп сконструйовано в 1934 р. німецьким вченим Є.Руска. Електронна мікроскопія значно поглибила уявлення про найтоншу будову системи органів тварин рослин, підтвердила положення клітинної теорії про єдність їх будови. Завдяки електронній мікроскопії стало очевидним, що клітина, крім ядра та цитоплазми, містить комплекс ще менших структур, і що в цілому вміст будь-якої клітини являє собою складну систему мембран, філаментів тощо.
Розвиток морфології та її розділів — цитології науки про будову та функціональне значення клітини, ембріології — яка вивчає закони генезу зародка і процес його розвитку, вчення про походження та функціональне значення тканин — власне гістології, та спеціальної гістології вчення про розвиток, будову та функціональне значення органів і їх систем стало можливим завдяки розвитку фізики і хімії.
В навчальному плані спеціальності 6.010200 «Фізична реабілітація» зазначені розділи об'єднані в одну дисципліну гістологія.
2. Методи гістологічного дослідження.
Розвиток гістології тісно пов'язаний з удосконаленням мікроскопів та розробкою методів мікроскопічного дослідження. Сучасна гістологія має різноманітні методи дослідження, які дають змогу всебічно вивчати розвиток, будову та функцію клітин, тканин, органів. Основним об'єктом дослідження при цьому є гістологічні препарати, виготовлені із фіксованих структур. Цей метод називають ще методом класичної гістології. Препарат може являти собою мазок (мазок крові, кісткового мозку), відбиток (печінки, селезінки, тимуса тощо), плівки (плеври, очеревини, м'якої мозкової оболонки), тотальні препарати (зародки на ранніх стадіях розвитку, статеві клітини). Постійні гістологічні препарати використовують у навчальному процесі й наукових дослідженнях.
Процес виготовлення гістологічного препарату полягає в наступному.
Першим етапом у ньому є одержання матеріалу. При цьому шматочки розміром близько 1x1 см3 вирізують гострою бритвою, не травмуючи об'єкту, останній повинен бути свіжим, брати його слід одразу ж після забою експериментальної тварини.
Другим етапом цього процесу є фіксація матеріалу. Її здійснюють відразу ж після вирізування шматочка. Полягає вона в зануренні його в фіксуючу рідину. Метою фіксації є збереження гістологічних структур. Фіксатором може бути 5— 10 % розчин формаліну: він швидко проникає у тканини, добре їх фіксує, легко видаляється після промивання у воді.
Фіксаторами є оцтова, пікринова, осмієва кислоти, нейтральний 10% формалін, етиловий та метиловий спирт. При необхідності застосовують різні складні фіксуючі суміші, які містять названі компоненти у різних співвідношеннях.
Третій етап виготовлення гістологічного препарату — зневоднення фіксованого матеріалу. Для цього використовують спирти зростаючої концентрації (від 50 до 100 градусів). Після зневоднення матеріал ущільнюється. Його здійснюють у насиченому розчині рідкого парафіну в ксилол при температурі 37° С, а потім в рідкому парафіні при температурі 55° С. У парафіні об'єкт просочується, йому дають змогу затвердіти при кімнатній температурі разом з парафіном у спеціальній формочці. Блок для електронної мікроскопії одержують ущільненням об'єкту в органічних смолах. Зрізи виготовляють на спеціальних приладах-мікротомах (для світлової мікроскопії тонкі зрізи товщиною 8 мкм, напівтонкі 0,5—1 мкм); для електронної мікроскопії ультратонк зрізи — 0,05—0,2 мкм виготовляють на ультрамікротомах.
Забарвлюють зрізи для збільшення контрастност зображення окремих структур при розгляді їх у мікроскопі. Методи забарвлення гістологічних структур різноманітні. Вибір їх залежить від мети дослідження. Гістологічні барвники за походженням поділяють на кислі, основні та нейтральні. Кислий фарбник — еозин забарвлює цитоплазму в рожево-жовтий колір; це синтетичний фарбник. Структури, що фарбуються кислими фарбниками, називають оксифільними. Основні фарбники забарвляюють ядра клітин і тому їх називають ядерними. Прикладом є гематоксилін — фарбник рослинного походження. Він фарбує ядро клітини в синьо-фіолетовий колір. Гістологічн структури, що здатні забарвлюватися основними барвниками, називають базофільними. Структури, що
сприймають кислі та основні барвники, нейтральними. Вони утворюються при сполученні водних розчинів кислого і основного барвників.
У гістологічній техніці знаходять широке використання спеціальні барвники. За їх допомогою фарбують речовини певної хімічної природи. Наприклад, альціановий синій використовують для визначення кислих глікозаміногліканів. Судан III забарвлю нейтральні жирові речовини в оранжевий колір, судан чорний В — ліпіди в чорний колір. Для забарвлення нервової тканини успішно користуються методикою імпрегнації азотнокислим сріблом тощо.
Після фарбування зрізи відмивають від залишку фарбника, зневоднюють етиловим спиртом, просвітлюють ксилолом, потім вміщують в тонкий шар бальзаму між предметним та покривним скельцями. Бальзам і скельця мають майже однаковий показник заломлення світла, що запобігає розсіюванню променів при проходженні їх через товщу препарату. Основний недолік цього класичного способу виготовлення препарату — поява штучного утворення — артефакту, що може бути причиною одержання негативних результатів. Однак, знаючи закономірності фіксування та зневоднення, артефактів можна уникнути. Так, якщо матеріал довго зберігати у формаліні, у ньому можуть утворитися темні пігментні зерна. Щоб запобігти їх появі, препарат ретельно промивають у проточній воді. Інша справа, коли порушують правила виготовлення препарату, з'являються волокна тканини, якою протирають скельця, пухирці повітря при накриванні препарату покривним скельцем, осад фарб, зазубрини мікротомного ножа, складки зрізу.
Крім основного класичного методу, в гістолог снує багато інших, які застосовують залежно від мети дослідження. Зокрема це такі методи:
флуоресцентна мікроскопія, яка дає змогу вивчити як власну (первинну) флуоресценцію речовин, так і вторинну, викликану фарбуванням клітинних структур спеціальними барвниками-флуорохромами. Останні, при взаємодії з різними компонентами клітини, утворюють специфічне світіння відповідних структур. Так, флуорохром — акридиновий оранжевий з ДНК дає зелене світіння, а з РНК — червоне.
Метод темнопольової мікроскопії полягає в тому, що дрібні часточки, які лежать за межами дозволеної здатності мікроскопа, стають видимими в променях, що йдуть під таким великим кутом і в об'єктив вони безпосередньо не потрапляють. В об'єктив потрапляє лише світло, відбите від цих часточок, вони мають вигляд світлих цяточок по темному фоні. Цей метод є цінним при вивченні живих колоїдів клітини. Є інші, широко використовуван методи: гістохімічний, ауторадіографічний, імуногістохімічний.
Мікрургія клітини і фракціонування клітинних стуктур. При вивченні властивостей живої клітини значне місце належить так званій мікрургії, яка дає змогу за допомогою спеціальних мікроманіпуляторів здійснювати операції на клітині. Із застосуванням мікрургії вивчають реакції на пошкодження і вилучення різних складових частин — ядра, ядерця, окремих хромосом.
Різновиди мікрургії вивчають локальний вплив на окремі частини хромосом вузького пучка променів (-гамма-променів, протонів, ультрафіолетових).
В цитології вивчають хімічний склад і властивості ізольованих структур та органоїдів клітин. Ізоляції їх досягають шляхом подрібнення тканин у гомогенізаторах, при цьому руйнуються клітинні оболонки. На фракції гомогенат поділяється в результаті центрифугування. Опрацьован засоби центрифугування гомогенатів у ступінчастому градієнті щільності. При цьому у пробірці нашаровуються розчини сахарози, що спадають від дна щільності, останнім нашаровується гомогенат.
Центрифугування вмісту такої пробірки да змогу одержати різні фракції по її вертикалі від найважчих (ядра, ядерця), які опускаються на дно і найлегших, як розміщуються ближче до поверхні (рибосоми, хромосоми, лізосоми).
Цитоспектрофотометрія да змогу визначити кількісний вміст речовин у клітин та їх складових елементів по поглинанню ними світлових променів певної довжини хвилі.
Авторадіографія. За цим методом аналізують розміщення у клітинах і тканинах речовин, які помічено радіоактивними ізотопами. Методом авторадіографії виявляють місця синтезу певних речовин, склад білків, шляхи внутрішньоклітинного транспорту. Ізотопи, введені в клітини, відновлюють бромисте срібло фотоемульсії, що покриває зріз. Після проявлення фотоемульсії помітні зерна срібла (треки), що свідчить про розміщення в клітинах мічених речовин.
Імуноцитохімічний метод дослідження. Для вивчення деяких складових білкового обміну користуються здатністю високомолекулярних речовин-антигенів викликати в клітинах утворення захисних глобулінів (антитіл) з'єднуватися з ними в комплекси. Приєднання до одного глобуліну флуоресціюючо речовини дає змогу виявити локалізацію іншого.
Гістохімічні методи дослідження. З їх допомогою виявляють хімічну природу складових елементів клітин і міжклітинної речовини, тканин тваринного організму. В основі гістохімічних методів застосовують специфічні хімічні реакції. За їх допомогою виявляють нуклеїнові кислоти, білки, амінокислоти, ліпіди, вуглеводи, ферменти.
Електронна мікроскопія да змогу виявляти субмікроскопічну будову клітин. При електронній мікроскопії використовують потік електронів, джерелом яких є розжарена вольфрамова нитка-катод. Під впливом підвищеної напруги в 80 кВт електрони набувають великої швидкості і спрямовуються до аноду, в центрі якого є отвір, через який вони проходять. В сучасних трансмісійних (просвічуючих) електронних мікроскопах роздільна здатність становить 0,1—0,7 нм. Метод скануючої електронної мікроскопії забезпечує об'ємне вивчення поверхонь об'єктів дослідження.
Прижиттєве дослідження тканин. Живі тканини культивують за межами організму. Шматочки тканини об'ємом до 1 мм2, одержані стерильно при мінімальному пошкодженні, розміщують у спеціальну камеру на слюду чи покривне скельце, де міститься штучне живильне середовище з відповідною температурою. Клітини в складі тканинних культур, особливо ембріональних, зберігають життя, діляться, здатні до гістологічної диференціації.
Значно поширений спосіб одношарових культур, у якому використовують клітини, одержані при подрібненні тканини дією трипсину. Метод прижиттєвого дослідження тканин дає змогу простежити рух клітин, їх поділ, ріст, реактивні зміни на дію різних факторів. З цією метою застосовують уповільнене фотографування.
3. Основи цитології. Історія розвитку.
Цитологія (від гр. цитос — комірка, клітина) — наука про походження, будову та функціональне значення елементарних живих систем організму. Термін «клітина» вперше застосував англійський фізик Р.Гук (1665), який за допомогою збільшуваних лінз розглядав зрізи пробки, серцевини бузини, а також стебла та корені різних рослин. У рослинній речовині Р.Гук називав клітинами правильно розміщені пустоти. М.Мальпігі (1671), Н.Грю (1671) підтвердили спостереження Р.Гука і назвали ці утворення «мішечками», «міхурцями». Н.Грю вважав, що ці «міхурці» об'єднуються в структури, як нагадують текстильні утворення і назвав їх «тканинами».
При всій невідповідності назв об'єктів, як називали термінами «клітина» та «тканина» вони збереглися й до цього часу. Пізніше А.Левенгук (1677—1680) — мистецький шліфувальник лінз, спостерігав одноклітинні організми вперше побачив еритроцити та спермії тварин. Мікроскопісти XVII століття, які спостерігали вперше клітини ссавців, не зіставляли їх з клітинами рослин. В середині XVII століття, тобто майже через століття після Мальпігі та Грю, досягнення оптичної техніки були такі незначні, що спостереження та рисунки мікроскопістів того часу мало відрізняються від описування і зображень, зроблених у XVII столітті.
Початок успішного вивчення клітин пов'язаний з розвитком мікроскопування. В кінці XVIII на початку XIX століття були створені ахроматичні мікроскопи, завдяки яким стали достовірнішими мікроскопічн спостереження, що дало змогу здійснити систематичне вивчення структурних елементів різноманітних тваринних та рослинних організмів. На той час змінилася уява про будову клітин, основним в організації клітини стали вважати не клітинну стінку, а її вміст — протоплазму. Поступово збагачувався матеріал про мікроскопічну організацію тварин і рослин та будову клітин (cellula), названих так ще Р. Гуком.
У той час незалежно один від другого рядом дослідників в клітинах рослин і тварин були відкрит ядра, що створило необхідну передумову для висунутого в 1837р. Пуркіньє положення про подібність в будові тваринних і рослинних клітин.
У 1838—1839 рр. Т.Шванн узагальнив ус попередні мікроскопічні дослідження і сформулював клітинну теорію. Він розглядав клітину як універсальний структурний компонент тваринних і рослинних організмів. У книзі «Мікроскопічні дослідження про відповідності в структур рості тварин і рослин» показано, що клітинна теорія стала найважливішою подією в біології, одним із вирішальних доказів єдності походження усієї живо природи. Вона значно вплинула на розвиток біології, медицини, інших наук. Основний зміст та значення клітинної теорії наведені в попередньому пункті.
Великий внесок у вивчення клітини і розвиток клітинної теорії було зроблено в середині XIX століття реформатором наукової і практичної медицини патологом Р.Вірховим (1858), який довів, що основою таких процесів як запалення, дистрофії, новоутворення та інші є ті чи нші зміни в клітинах, і обґрунтував нові напрями досліджень: целюлярну фізіологію та целюлярну патологію.
Розвиваючи дані своїх попередників про спадкоємність розмноження клітин шляхом їх поділу Р.Вірхов висунув відому тезу: «Кожна клітина від клітини» — «Omnis cellula e cellula », яка не втратила свого значення і нині, але була в подальшому обґрунтована на молекулярному рівні.
В другій половині XIX та на початку XX століття проводили дослідження тонкої будови клітини. В той період виникли гіпотези будови протоплазми (фі-брилярна — В.Флемінга, гранулярна — Р.Алтман, комірчаста О.Бючлі). Однак усі ці гіпотези мали один недолік — вони розглядали протоплазму в стані стабільної структури. В той же час були відкрит закономірності поділу клітин (Є.Руссов (1872), А.Шнейдер (1873), І.Д.Чистяков (1874), Є Страсбургер (1875), П.І.Перемежко (1878), В.Флемінг (1878), Бовері (1879) та інш.). Описана центросома (Бовер (1875)), мітохондрії (Бенда (1897)), комплекс Гольджі (К.Голь-дж (1898)).
У XX столітті завдяки електромікроскопічним дослідженням, одержані дані про ультраструктурну організацію клітини.
Вчення про клітину має велике значення для прогресивної еволюції живих істот; воно пояснює точність передачі спадково нформації від організму до організму у форму, що забезпечує виникнення і збереження нових властивостей, утворення тканин, що забезпечують здійснення основних функцій організму являють собою будівельний матеріал для його органів, збільшення росту організму, що забезпечує йому високий енергетичний потенціал, чим створює сприятливі умови в складному взаємозв'язку із зовнішнім середовищем, а також зміну зношуваних в процесі життєдіяльності структурних елементів при фізіологічній регенерації і ушкоджених частин тіла (репаративна регенерація).
Нині в цитології існують такі напрями, як цитоморфологія — наука про будову клітин, цитофізіологія — наука про функціональні прояви клітин, цитохімія — наука про хімічний склад клітин, цитогенетика — наука про спадковість та мінливість клітин, цитопатологія — наука про патологічні зміни клітин.
Сучасну цитологію на основі експериментальних досліджень застосовують при цитодіагностиці хвороб, цитоімунологічних пробах, у тканевоспецифічних регуляціях розмноження клітин, реактивних змін під впливом факторів зовнішнього середовища тощо.
Науково-технічний прогрес, успіхи розвитку методів дослідження дали змогу визначити ультраструктурну організацію клітини та неклітинних структур, зрозуміти процеси диференціювання, регенерації, передачі спадкових ознак тощо.
4. Біологія клітин: органели, їх будова і функції.
Серед живих організмів існує два типи організації клітин: прокаріотичні та еукаріотичні. До прокаріотичних відносять клітини бактерій та синьозелених водоростей, їх називають доядерними клітинами, вони обмежені плазматичною мембраною, зовні від плазматично мембрани знаходиться клітинна стінка, яка похідним клітинної активності. Прокаріотичні клітини не мають морфологічного ядра, але містять нуклеоідну зону, заповнену ДНК, а в матриксі цитоплазми прокаріотів розміщуються рибосоми.
Еукаріотичн клітини належать до вищого типу, їх обов'язковою структурою є: цитоплазма, клітинна оболонка (плазмолема) та клітинне ядро, яке відокремлене від цитоплазми ядерною мембраною. У клітинах еукаріотичного типу в цитоплазмі є спеціальні структури — органели, які виконують окремі специфічні функції. До органел відносять мембранні та немембранні структури (рис. 1).
У тварин найменшою одиницею живого є клітина еукаріотичного типу.
Узагальнюючи уяву про будову клітин, як одиниць живого, їх розмноження і ролі у формуванні багатоклітинних організмів, клітина являє собою елементарну саморегулюючу живу систему, основу будови, розвитку та життєдіяльності усіх тваринних і рослинних організмів.
Виходячи із положення клітинної теор Т.Шванна, клітина є частина цілісного організму, найменша його одиниця, їй притаманні усі властивості, що відповідають визначенню поняття «живого»; клітини рослин і тварин мають однаковий загальний план будови подібність загально-клітинних функцій для підтримання х життя та розмноження, яке відбувається шляхом поділу. У багатоклітинному організмі клітини мають спеціалізацію будови і виконують різні функції. Однак вони підпорядковані цілому, входять до складу тканин, органів, систем органів, пов'язані міжклітинними та нейрогуморальними формами регуляції. Величезна різноманітність живої матерії та ндивідуальність, що притаманні кожному організмові, різноманітність форм та функцій клітин у багатоклітинному організмі пов'язані з білками та нуклеїновими кислотами. Нуклеїнові кислоти забезпечують передачу спадкової інформації, відіграють велику роль у процесах біосинтезу білків. Білки беруть безпосередню участь у всіх реакціях, що відбуваються в клітинах та в організмі, характеризуються високим ступенем специфічності. Білкову природу мають усі ферменти, антитіла, деякі гормони тощо. До складу живої матерії входять вода, вуглеводи, ліпіди, вітаміни, мінеральні солі. Щодо фізико-хімічного стану клітина являє собою колоїдну систему, залежно від функціонального стану здатну переходити із рідкого в гелепо-дібний та в зворотній стан. Детальна інформація про фізико-хімічні особливості речовин, що входять до складу живої системи, вивчає наука біохімія.
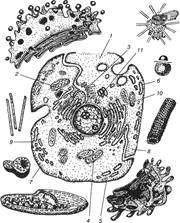
рис.1. Загальна схема будови клітини.
1 — плазмолема; 2 – комплекс Гольджі; 3 — цитоплазма; 4 — мітохондрія;
5 – ендоплазматична сітка; 6 - рибосома; 7- лізосома; 8 - мікротрубочка;'
9 — мікрофіламенти; 10 - ядро; 11- центросома.
Структурн компоненти клітин.
Цитоплазма (cytoplasma), від навколишнього середовища відокремлена пла-змолемою; до неї належать гіалоплазма, органели, включення.
Плазмолема різновидом мембран клітини. Це тонкі (6—10 нм) пласти ліпо- протеїдної природи, до складу яких входять 40% ліпідів, близько 60% білків, значна частина клітинних мембран містить 5—10% вуглеводів. Серед ліпідів знаходяться органічні речовини, як гідрофобними — погано розчиняються у воді та характеризуються значною розчинністю в органічних розчинниках та жирах ліпофільні. Серед ліпідів, що входять до складу клітинних мембран, знаходяться фосфоліпіди, сфінгомієліни, холестерин. Особливістю ліпідних мембран поділ молекул на дві функціонально різні частини: гідрофобні неполярні, що не несуть зарядів, «хвости», складаються із жирних кислот, та гідрофільні, заряджені полярні «головки». Це визначає здатність ліпідів довільно утворювати двошарові (біліпідні) мембранні структури товщиною 5—7 нм. Клітинні мембрани можуть значно різнитися між собою за ліпідним складом та білковими молекулами. Багато мембранних білків складаються з двох частин, з ділянок багатих полярними (що несуть заряд) амінокислотами, та ділянок, збагачених неполярними амінокислотами. Такі білки в ліпідних шарах мембран розміщуються так, що їх неполярн ділянки ніби занурені в «жирну» частину мембрани, де знаходяться гідрофобні ділянки ліпідів. Полярна (гідрофільна) частина цих білків взаємодіє з головками ліпідів і оберальних білків, існують білки частково вмонтовані у мембрану — напівінтегральні та примембранні, вмонтовані в біліпідний шар. За біологічним значенням білки мембран поділяються на білки-ферменти, переносники, рецепторні та структурні.
Вуглеводи мембран входять до складу не у вільному стані, а у зв'язку з молекулами ліпідів або білків. Такі речовини називають відповідно гліколіпідами та глікопротеїдами. На поверхн плазмолеми вони формують надмембранну зону — глікокалікс, товщиною 3— 4 нм. За його участю здійснюється взаєморозпізнання клітин та взаємодія їх між собою. Вс мембрани є бар'єрними структурами.
Плазмолема викону функції розмежування цитоплазми із зовнішнім середовищем, рецепції та транспорту різних речовин, серед яких важливе місце належить забезпеченню оптимального рівня асиметрії концентрації іонів натрію і калію у клітині та за її межами, а також забезпечує взаєморозпізнавання і взаємодію клітин з утворенням міжклітинних контактів, формування структури клітинної поверхні, рецепцію сигналізації з боку зовнішнього середовища.
Транспортна функція плазмолеми здійснюється шляхом дифузії пасивного перенесення різних речовин — води, іонів, деяких низькомолекулярних сполук. Інші речовини проникають через мембрану шляхом активного перенесення проти градієнта концентрац за рахунок розщеплення АТФ. Так транспортуються цукри, амінокислоти. Ц процеси відбуваються за участю спеціальних білків — переносників. Транспорт із зовнішнього середовища в клітину називають ендоцитозом, за межі клітини — екзоцитозом. Великі молекули та х агрегати проникають у клітину шляхом фагоцитозу, який був вперше описаний І.І. Меч-никовим.
Поглинання частинок рідини називають піноцитозом. Загальним для цих процесів є те, що поглинання на поверхн плазмолеми відбувається шляхом оточення речовини ділянкою плазматичної мембрани у вигляді вакуолі, яка переміщується у цитоплазму. Поглинут частинки розщеплюються за допомогою ферментів і їхні складові засвоюються клітиною. Таким чином, поглинуті речовини у середині мембранних вакуолей, утворених із елементів плазмолеми, підлягають внутрішньому клітинному травленню. Процес, при якому поглинені частинки в оточенні мембрани проходять через цитоплазму і виводяться без змін за меж клітини називають цитопемпсисом.
Плазмолема бере участь у виведенні речовин із клітини (екзоцитоз). У цьому випадку внутрішньоклітинні продукти (білки, мукополісахариди, жирові краплі тощо) містяться у вакуолі або міхурці і відмежовані від гіалоплазми мембраною наближаються до плазмолеми. У місцях контакту плазмолема та мембрана вакуолі зливаються і вміст вакуолі виходить за межі клітини.
Екзоцитоз поділяють на такі різновиди: секрецію — виділення клітиною продуктів її синтетичної діяльності; екскреція виділення шкідливих продуктів метаболізму; клазматоз — видалення за межі клітин окремих її структурних компонентів.
Ендоцитоз та екзоцитоз відбуваються за участю пов'язаних з плазмолемою системи фібрилярних компонентів цитоплазми — мікротрубочок та скорочувальних мікрофіламентів, вони з'єднуються у певних ділянках плазмолеми і утворюють кортикальний шар.
Рецепторні функції плазмолеми пов'язані з локалізацією на ній спеціальних структур, які беруть участь в специфічному пізнанні хімічних та фізичних факторів. Клітинна поверхня ма велику кількість рецепторів, що визначають можливість специфічних реакцій. Існують рецептори до біологічно активних речовин — гормонів, медіаторів, антигенів тощо. Складні процеси рецепц основою взаєморозпізнання клітин, важливою і необхідною умовою снування багатоклітинних організмів.
Міжклітинн контакти
Сполучення між клітинами у складі тканин та органів багатоклітинних організмів утворені складними спеціальними структурами, які називають міжклітинними контактами. Розрізняють такі типи міжклітинних контактів. Просте міжклітинне сполучення — плазмолеми сусідніх клітин наближаються одна до одної на відстань 15—20 нм. При цьому відбувається взаємодія шарів глікокалікса. Розрізняють також щільне з'єднання за типом замка, при цьому шари двох плазмолем наближен максимально — ділянки плазмолем двох сусідніх клітин ніби зливаються. Ця область непроникна для молекул та іонів. Тип з'єднання плазмолем називають десмосомою. Вона являє собою площу діаметром до 0,5 мкм, інколи має вигляд шарів; між мембранами знаходиться зона з високою електронною щільністю; зміцнення зв'язку між клітинами досягається за допомогою фібрилярних структур цитоплазми та кортикального шару плазмолеми. Спостерігаються з'єднання типу напівдесмосом, якщо десмосома складається лише з однієї пластинки прикріплення.
Щільний замикаючий контакт характерний для апікальної поверхні клітин каймистого епітелію, а також ендотелію, мезотелію. Такий контакт характеризується максимальним зближенням плазматичних мембран сусідніх клітин, проміжок ущільнюється за рахунок анастомозуючих фібрил та іонів кальцію. При цьому зовнішні гідрофільні шари і глікокалікс суміжних плазмолем тісно зливаються в один суцільний шар завтовшки 2—3 нм. У серцевих м'язах зустрічаються щілинні контакти (нексуси), що забезпечують безпосередній обмін речовин між сусідніми клітинами. Так відбувається перенесення іонів та дрібних молекул.
Синаптичні з'єднання характерні для нервово тканини, або між нервовою клітиною і м'язом, у ділянці якого відбувається передача імпульсу.
Синапс спеціалізований контакт, що забезпечує передачу нервового збудження. До його складу входять: ділянка плазмолеми пресинаптичної мембрани відростка нервової клітини, з якої відходить імпульс, ділянка плазмолеми постсинаптичної мембрани клітини, яка сприймає сигнал. До сигналу входить синаптична щілина, що розмежовує пресинаптичну та постсинаптичну мембрани і, заповнені нейромедіатором, синаптичн пухирці. Синапси забезпечують однобічну передачу інформації від клітини до клітини завдяки медіатору.
Цитоплазма.
До складу цитоплазми (Cytoplasma) входять 1) гіалоплазма, 2) органели та 3) включення.
1) Гіалоплазма — це прозора основна плазма, або матрикс цитоплазми, що позначає її внутрішн середовище. У електронному мікроскопі матрикс цитоплазми має вигляд гомогенної та тонкозернисто речовини з низькою електронною щільністю. Гіалоплазма являє собою складну колоїдну систему і містить різні біополімери: білки, нуклеїнові кислоти, полісахариди тощо. Ця система здатна переходити із рідкого стану в гель навпаки. Окремі зони гіалоплазми можуть змінювати свій агрегатний стан залежно від умов або функціонального завдання. Деякі молекули білків — тубулінів можуть бути диспергійовані в гіалоплазмі, але в певні моменти вони починають збиратися і утворювати мікротру-бочки. Таким же чином, у гіалоплазмі можуть виникати і розпадатися різноманітні фібрилярні, нитчасті комплекси білкових молекул. До ферментів матрикса відносять ферменти гліколізу, метаболізму цукрів, азотистих сполук, амінокислот, ліпідів та інших важливих сполук. У гіалоплазмі відбувається постійний рух іонів до плазматичної мембрани і від неї, до мітохондрій, ядра, вакуолі. Гіалоплазма є зоною переміщень молекул АТФ. В гіалоплазмі містяться органели та включення.
2) Органели постійні мікроскопічні та ультраструктурні утворення, що виконують життєвоважливі функції клітин. Серед органел розрізняють мембранн органели — мітохондрії, ендоплазматичну сітку, лізосоми, пероксисоми, комплекс Гольджі; до немембранних органел відносяться — рибосоми, мікрофіла-менти, мікротрубочки, центросома. Всі вони належать до органел загального призначення. Поряд з цим в деяких клітинах існують спеціальн органели — тонофібрили епітеліальних клітин, міофібрили міоцитів та м'язових волокон, нейрофібрили нервових клітин.
Мембранні органели
Мітохондрії. Термін «мітохондрія» введено Бендою у 1897р. В світловому мікроскоп мітохондрії мають вигляд круглих зерен та коротких паличок. При розгляді під електронним мікроскопом кожна мітохондрія має овальну або видовжену форму. У них розрізняють зовнішню гладеньку та внутрішню мембрани, від останньо всередину мітохондрії відходять вирости — кристи. Проміжки між кристами заповнені матриксом — електронно-щільною речовиною, у якій виявляють ниткоподібні утворення товщиною 2—3 мм та гранули товщиною 15—20 мм. Ниткоподібні утворення матрикса являють собою молекули ДНК і РНК, а гранули мітохондріальні рибосоми. У матриксі і у внутрішній мембрані містяться білки — ферменти, що забезпечують синтез АТФ шляхом окисного фосфорилювання аденозиндифосфату. Середній розмір сильно варіює і становить понад 20% загального об'єму цитоплазми і містить до 35% загальної кількості білка в клітині. Мітохондрії концентруються у тих місцях цитоплазми, де є потреба в АТФ. Основною функцією їх є синтез АТФ, який відбувається в результаті процесів окислення органічних субстратів і фосфорилювання АДФ.
Мітохондрії також беруть участь у регуляц обміну води, депонування іонів кальцію, продукції попередників стероїдних гормонів. Мітохондрії здатні рухатися; їх кількість збільшується шляхом поділу, або брунькуванням початкових мітохондрій. Мітохондрії чутливі до різних зовнішніх впливів: голодування, дії рентгенівських променів, наркотиків тощо.
Ендоплазматична сітка. Її вперше описав К.Портер у 1945 р. Вона являє собою субмікроскопічну органелу, яка утворює внутрішньоцитоплазматичну циркуляційну систему. Вона є замкненою сукупністю канальців, мішечків та цистерн, утворених безперервною біомембраною. Розрізняють два типи гранулярну та агранулярну. Ширина канальців гранулярної сітки від 20 до 1000 нм, з боку гіалоплазми її мембрани покриті рибосомами. Останні беруть участь у синтезі білків на «експорт» і білків-ферментів, а також білків, що нагромаджуються в цистернах ендоплазматичної сітки та здатн транспортуватися у вакуолі комплексу Гольджі, де перетворюються і входять до складу лізосом або секреторних гранул, утримання яких залишається ізольованим від гіалоплазми мембраною. У ряді випадків у канальцях або вакуолях гранулярної ендоплазматичної сітки відбувається модифікація білків, зв'язування їх з цукрами утворення секреторних гранул. Гранулярна ендоплазматична сітка здатна синтезувати мембранні інтегральні білки, які вмонтовуються в товщу мембран.
На відміну від гранулярної ендоплазматично сітки на мембранах агранулярної ендоплазматичної сітки відсутні рибосоми. Діаметр її канальців і міхурців 50— 100 нм. Функція гладкої ендоплазматичної сітки пов'язана з метаболізмом ліпідів, синтезом глікогену, депонуванням іонів кальцію, дезактивацією отруйних речовин, що особливо характерно для гепатоцитів. Мембрана ендоплазматичної сітки безпосередньо контактує з плазмолемою клітин та мембранами.
Лізосоми — мембранні органели. Вони являють собою кулясті структури розміром 0,2— 0,4 мкм, містять понад 60 гідролітичних ферментів, здатних до розщеплення біополімерів різної хімічної природи. Із ферментів лізосоми містять протеїнази, нуклеази, глюкозидази, фосфатази, ліпази. Ферменти локалізуються у лізосомах і утримуються у них за допомогою ліпопротеїново мембрани, яка обмежує і відокремлю х вміст від зовнішніх субстратів. Основна фізіологічна функція лізосом внутрішньоклітинне травлення в зв'язку з процесами фагоцитозу, піноцитозу тощо. Залежно від ультраструктурних та функціональних особливостей лізосоми поділяють на первинні, їх ферменти знаходяться у неактивному стані; вторинні, або фагосоми (активовані ферменти в них безпосередньо контактують з розщеплюваними біополімерами), а також залишков тільця, оточені біомембраною нерозщеплені залишки. Поряд з цим лізосоми можуть брати участь у розщепленні власних макромолекулярних комплексів клітин аутофагоцитозу.
Пероксисоми субмікроскопічні мембранні органели. Відіграють вирішальну роль у процесах детоксикації клітини. Пероксисоми — тільця розміром 0,3-1,5 мкм, обмежені мембраною, містять гранулярний матрикс, в центрі якого розміщуються кристолоїдноподібн структури із фібрил і трубок. Ферментні системи пероксисом (каталаза) спрямовані на утилізацію хімічно активного атомарного кисню, а також забезпечують розщеплення етилового спирту, сечової кислоти, регуляцію обміну ліпідів.
Комплекс Гольджі. Перш відомост про комплекс Гольджі належить К.Гольджі, який в 1898р. описав цю органелу у склад нервових клітин. Під світловим мікроскопом комплекс Гольджі має вигляд сітчастого утворення. Під електронним мікроскопом він являє собою мембранні структури, що мають вигляд цистерн товщиною до 25 нм, сплющених у центральній частині і розширених на периферії. Окрему зону скупчення цих мембран називають діктіосомою. В проміжках між окремими цистернами знаходяться тонкі прошарки гіалоплазми. У комплексі Гольджі завершується процес формування продуктів синтетичної діяльності клітини кінцеве глікозування. Комплекс Гольдж бере участь у сегрегації і нагромадженні продуктів, синтезованих в цитоплазматичному ре-тикулумі, у їх хімічній трансформації. У цистернах комплексу Гольдж синтезуються полісахариди, які комплексуються з білками, що сприяє утворенню мукопротеїдів, а також виведенню готових секретів за межі клітини. Крім того, комплекс Гольджі забезпечу формування клітинних лізосом.
Не мембранні органели
Рибосоми. За розміром рибосоми становлять 25x20x20 нм. До їх складу входять велика і мала субодиниці рибосомної РНК та білок.
Для стабілізації структури рибосом необхідн они магнію. Кожна із субоди-ниць побудована з нуклеопротеїдного тяжа, де РНК взаємодіє з різними білками утворює тіло рибосоми, у яких амінокислоти сполучаються, тобто синтезуються білкові молекули. Кілька рибосом, з'єднаних спільною ниткою інформаційної РНК, називають полісомами. Останні полісоми пов'язані з мембранами ендоплазматичної сітки, синтезують білки для виведення за межі клітини. Ступінь інтенсивності синтетичної діяльності вільних рибосом менший, а утворені білки використовуються для внутрішніх потреб клітини.
Мікрофіламенти. Це тонкі волоконця діаметром до 5 нм, побудовані з різних білків — актину, міозину та ін. Вони розміщені переважно в кортикальній зоні клітини у складі її цитоплазматичних виростів і виконують роль цитоскелету, а також скоротливого апарату клітини.
Мікротрубочки являють собою прямі, довг порожнисті білкової природи циліндри. їх зовнішній діаметр досягає 25 нм, а внутрішній просвіт — 15 нм; містять глобулярний білок тубулін. Мікротрубочки входять до складу центросоми та базальних тілець, а також є основними структурними елементами війок та джгутиків. Мікротрубочки забезпечують рухливість клітинних органел, а також беруть участь в утворенні цитоскелету.
Центросома. Описана В.Флемінгом у 1875р. Міститься вона у клітинах тварин, за винятком яйцеклітин. Центросома складається з двох центріолей, оточених центросферою, що являє собою позбавлену органел гіалоплазму, яку радіально пронизують мікрофіламенти і мікротрубочки. Основою будови центріолей є дев'ять триплетів паралельно розміщених мікротрубочок, які формують циліндр 200x500 нм. Крім мікротрубочок, до складу центріолі входять так зван «ручки», за їх допомогою триплети пов'язані між собою. У склад «ручок» міститься білок тубулін, що має АТФ-азну активність і якому належить важлива роль у механізмі рухових функції центріолей. Довг осі обох центріолей розміщені у взаємно перпендикулярних площинах. Перед поділом клітини центріолі подвоюються (настає дупликація) з подальшим розходженням кожної новоутворено пари до полюсів клітини. Дв розміщені поряд центріолі називають диплосомою. Вважається, що центріолі беруть участь в індукції полімерізації тубу-лінів при утворенні мікротрубочок. Центросома забезпечує розходження хромосом при поділі клітини.
Війки та джгутики. В світловому мікроскоп вони мають вигляд виростів клітини. В основі війок і джгутиків у цитоплазмі видно базальні тільця. Війки являють собою тонкі циліндричні вирости цитоплазми з діаметром 200 нм, які від основи до верхівки покрит плазмолемою. У середині виросту знаходиться аксонема, що складається із мікротрубочок. Базальне тільце занурене в цитоплазму; за своєю будовою воно подібне до центріолі, і структурно пов'язане з аксонемою у єдине ціле. У різних клітинах рух війок і джгутиків може нагадувати рух маятника, він хвилеподібний. Фіксовані клітини з війками на апікальній поверхні рухом війок можуть затримувати й транспортувати окремі частини, рідину.
3) Включення.
Це непостійні компоненти цитоплазми, їх форма не є суворо визначеною; включення тісно пов'язані з метаболічним станом клітин. Серед включень розрізняють: трофічні, пігментні, секреторні, екскреторні.
Трофічними включеннями є глікоген полісахарид, який нагромаджується у гіалоплазмі як резервний енергетичний матеріал м'язової тканини, гепатоцитах. До трофічних включень, що також беруть участь у щоденному метаболізмі клітин, належать крапельки нейтрального жиру жирових клітин рихлої сполучної тканини, в цитоплазмі яких знаходяться гігантські краплини нейтральних жирів. Ліпідні включення складніші за нейтральні жири. За хімічним складом вони постійні в клітинах наднирників, олігодендроглії тощо. Метаболізм їх спрямований на утворення специфічних продуктів ліпідної природи (гормони, мієлін). Білкові трофічні включення зустрічаються порівняно рідко. Прикладом їх можуть бути білков брилки в гепатоцитах; жовткові шари за х хімічним складом, крім білка, мають також інші речовини.
Пігментні включення. Серед них розрізняють ендогенні — гемоглобін, міоглобін, гемосидерин, білірубін, меланін, ліпофусцин та екзогенні — каротин тощо. Серед ендогенних пігментних включень важливе значення мають гемоглобін та міоглобін, які за хімічною будовою близькі між собою, здатні зв'язувати кисень і віддавати його при необхідності. Інші пігментні включення мають різну хімічну природу функціональне значення. Так, у тілі тварин, у клітинах є велика кількість включення бурого пігменту меланіну. Ці клітини називають меланоцитами. Вони нагромаджують меланін у вигляді овальних гранул меланосом. Меланін виконує захисну функцію проти дії ультрафіолетового випромінювання.
Секреторні включення характерні для залозистих клітин і являють собою або містять у собі продукти, що виділяються із клітин та мають специфічне значення для нормального функціонування організму. Такі властивості мають плазмоцити, що продукують захисний білок — гама-глобулін; клітини слинних залоз продукують різний за своїм складом секрет, який у своїй сукупності утворює слину. Слід зазначити, що в тваринному організмі значній кількості клітин характерні ознаки секреції, які відображають різновиди внутрішньоклітинного метаболізму.
Екскреторні включення це продукти метаболізму, які виділяються з клітини.
Ядро (nucleus). Ядро є однією з важливих складових частин клітини. Воно забезпечує збереження підтримку спадкової інформації у вигляді незмінної структури ДНК. В ядр відбувається відтворення або редуплікація молекул ДНК, що дає змогу при мітозі двом дочірнім клітинам одержувати цілком однакові в якісних і кількісних відношеннях об'єми генетичної інформації.
Другою групою клітинних процесів, що забезпечуються функцією ядра є утворення власне апарата білкового синтезу. Це не лише синтез, транскрипція на молекулах ДНК різних інформаційних РНК, а й транскрипція усіх видів транспортних і рибосомних РНК. У ядрі також утворюються субодиниці рибосом шляхом комплексування синтезованих в ядерці рибосомних РНК з рибо-сомними білками, які синтезуються в цитоплазмі і переносяться в ядро.
Ядро може перебувати у мітотичному стані — під час поділу клітини та в інтерфазному — між поділами — метаболічні ядра. У живій клітині інтерфазне ядро оптично пусте, в ньому видно лише ядерце. При дії різних пошкоджуючих агентів клітина набуває стану паранекрозу (межа між життям та смертю). З цього стану клітина може повернутися до нормальної життєдіяльності або загинути. В ядрі у цей час морфологічно розрізняють зміни, характерні для загибелі клітини: каріопікноз — ущільнення, каріорексис — розпад, каріолізис розчинення.
Таким чином, ядро являє собою одну з важливих складових частин клітини. Усі клітини тварин містять ядро, за винятком зрілих клітин крові — еритроцитів. Форма ядра різна, однак в більшості випадків вона відповідає формі клітини. Так, ядро лімфоцита кулясте, клітин гладеньких м'язів паличкоподібне, гепатоцитів — кулясте, ядро жирових клітин під впливом жирових накопичень сплющене та відтиснене до плазмолеми, у слинних залозах, клітини яких продукують слизовий секрет, ядро також сплющене і розміщується у базальній частині клітини. Ядро лейкоцитів, крім кулястого, може бути сегментованим, бобоподібним, паличкоподібним, мати вигляд підкови. Ядро та цитоплазма дина інтегрована система, що знаходиться у постійній рівновазі. Об'єм ядра і цитоплазми кожного типу клітин має сво постійне співвідношення. До складу ядра входять ядерна оболонка, хроматин, ядерце, каріоплазма.
Ядерна оболонка — нуклеолема складається із зовнішньої і внутрішньо біологічних мембран, відокремлених перинуклеарним простором шириною 20— 60 нм. Кожна з мембран має товщину до 8 нм і морфологічно подібна до інших клітинних мембран. Ядерна мембрана відокремлює вміст ядра від цитоплазми, зовнішня — безпосередньо контактує з цитоплазмою клітини, має ряд структурних особливостей, що дає змогу віднести до мембранної системи ендоплазматичної сітки. На зовнішній мембрані з боку гіалоплазми знаходяться полірибосоми, а сама зовнішня ядерна мембрана переходить в мембрани ендоплазматичної сітки. Внутрішня мембрана пов'язана з хромосомним матеріалом ядра.
Ядерна мембрана містить ядерні пори, утворен в результаті злиття зовнішньої та внутрішньої ядерних мембран. При цьому утворюються округлі наскрізні перфорації до 90 нм. Вони заповнені складноорганізованими глобулярними та фібрилярними структурами, які разом з мембранною перфорацією утворюють комплекс пори. Він побудований з трьох рядів гранул по вісім штук у кожному ряді, діаметр гранул 25 нм. Від гранул відходять фібрилярн відростки. Фібрили, що відходять від периферійних гранул, можуть сходитися в центрі і утворювати своєрідн перегородки поперек пори, так звану діафрагму пори. Розмір ядерних пор у кожного виду клітин є величиною сталою. Кількість ядерних пор залежить від метаболічної активності клітин, густина їх на поверхні нуклеолеми вище у клітин з високою метаболічною активністю.
Ядерна оболонка виконує бар'єрну функцію, відокремлює вміст ядра, його генетичний матеріал від цитоплазми, обмежує вільний доступ в ядро та вихід з нього різних речовин, регулює транспорт макромолекул між ядром і цитоплазмою. Ядерна оболонка бере участь у створенні внутрішньоядерного порядку шляхом фіксації хромосомного матеріалу в інтерфазі до внутрішньої ядерної мембрани.
Хроматин. Завдяки здатності сприймати фарбники, ця складова частина ядра названа «хроматин» (Флемінг, 1880). Здатність хроматину сприймати лужні фарбники свідчить про його кислотні властивості, які визначаються тим, що до складу хроматину входить ДНК у комплексі з білками. Властивостями фарбуватися утримувати ДНК володіють і хромосоми, що спостерігається під час мітотичного ділення клітин. Таким чином, хроматин нтерфазних ядер являє собою хромосоми, які у цей час втрачають свою компактну форму, розпушуються, деконденсуються. Зону повної деконденсації і її ділянки мають назву еухроматин. Під час мітозу весь еухроматин конденсується і входить до складу хромосом. При неповному розпушуванні хромосом в нтерфазному ядрі видно ділянки конденсованого хроматину, який називають гетерохроматином.
До складу хроматину входять складні комплекси дезоксирибонуклеопротеї-дів, що складаються з ДНК і спеціальних хромосомних білків — гістонів. Хроматин також містить РНК. В кількісному співвідношенні ДНК, білок та РНК становлять 1:1,3:0,2.
Фракція білків хроматину становить 60—70% сухої маси. До білків хроматину відносять так зван гістони та негістонові білки.
Гістони лужні білки, збагачені лізином і аргініном. Вони забезпечують специфічну укладку хромосомної ДНК і беруть участь у регуляції транскрипції.
Гістони розміщені вздовж молекул ДНК не рівномірно, а у вигляді блоків. У один такий блок входить вісім молекул гістонів, які утворюють нуклеосому, розмір яких 10 нм. При утворені нуклеосом відбувається компактизація, надспі-ралізація ДНК, що приводить до скорочення довжини хромосомної фібрили у п'ять разів. Саме хромосомна фібрила має вигляд нитки намиста. Такі фібрили додатково повздовжньо конденсуються і утворюють основну елементарну фібрилу хроматину товщиною 25 нм.
Негістонові білки інтерфазних ядер утворюють всередині ядра сітку, яку називають ядерною білковою матрицею. Вона являє собою основу, що визнача морфологію і метаболізм ядра.
Хроматин є структурним аналогом хромосом, як можна спостерігати під час поділу клітин. В деяких випадках ціла хромосома в період інтерфази може залишатися у конденсованому стані, вона має вигляд грудочки гетерохромати-ну. Наприклад, одна з Х-хромосом у соматичних клітинах самки залишається у конденсованому стані на стад дроблення зиготи. Вперше цей хроматин був досліджений у ядрі клітини кішки і описаний М.Барром і Л.Бертрамом у 1949 р. Його назвали статевим хроматином, або тільцем Барра. Визначення статевого хроматину використовують для встановлення генетично статі організму.
В ядрах, крім хроматинових ділянок матриксу, мають місце перихромати-нові фібрили, перихроматинов та інтерхроматинові гранули, які містять РНК. Матрицями для синтезу РНК є різн гени, розміщені по деконденсованих ділянках хроматинових фібрил. У хроматині міститься 1% ліпідів, роль їх не визначено.
Ядерце (nukleolus). В клітині міститься одне або кілька кулястої форми тілець розміром до 5 мкм. Вони добре фарбуються різними фарбниками. Ядерця багаті на РНК. Ядерце — найщільніша структура ядра, що є похідним хромосоми, одним з її локусів з найвищою концентрацією та активністю синтезу РНК в інтерфазі. Ядерце — це місце утворення рибосомних РНК та рибосом, на яких синтезуються поліпептидні ланцюги в цитоплазмі. Утворення ядерець та їх кількість пов'язані з активністю і кількістю ядерцевих організаторів. Навколо ядерця завжди виявляють зону конденсованого хроматину. ДНК ядерцевого організатора зображена численними копіями генів РНК: на кожному з цих генів синтезується високомолекулярний попередник РНК, який перетворюється у коротші молекули РНК, що входять до складу субодиниць рибосоми. Під електронним мікроскопом в ядерці виявляють гранули діаметром 15—20 нм і фібрили товщиною 6—8 нм. Гранулярний компонент має вигляд нитки, товщина нуклеолоне-ми 0,2 нм. Фібрилярний компонент ядерця рибонуклеопротеїнові тяжі — попередники рибосом, що дозрівають.
Каріоплазма — це рідка частина ядра, в якій містяться ядерн структури. Досліди показали, що після видалення з ядер хроматину і мембран, вони зберігають свою цілісність. Під електронним мікроскопом встановлено, що до складу каріошіазми входять комплекси пор з фібрилярним периферійним шаром, ядерцеві та численні фібрили, що знаходяться у міжхроматинових районах. Весь комплекс цих структур називають білковим ядерним матриксом. Компоненти матриксу являють собою динамічну структуру; ядерний матрикс відіграє важливу роль в підтримці загальної структури ядра, може брати участь в регуляції синтезу нуклеїнових кислот.
Література:
1. Гистология, цитология и эмбриология / Под ред. О.В.Волкова, Ю.К.Елецкого. - М: Медицина, 1996.
2. Луцик О.Д., Іванова А.Й., Кабак К.С., Чайковський Ю.Б. Гістологія людини. - К.: Книга плюс, 2003. - 3-тє видання. - 592 с.
3. Новак В.П., Пилипенко М.Ю., Бичков Ю.П. Цитологія, гістологія, ембріологія.: Підручник. – К.: ВІРА-Р, 2001. 288 с.
© 2010 Интернет База Рефератов