
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Курсовая работа: Физиология молокообразования
Курсовая работа: Физиология молокообразования
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РФ
Московский государственный университет прикладной биотехнологии
Кафедра анатомии, физиологии и животноводства
Курсовая работа:
Физиология молокообразования
Выполнила:
Студентка В-II-10
Васильева Ю.Н.
Проверила:
Преп. Рыжкова Е.М.
Москва 2004
Содержание
Введение
Состав молока. Биосинтез его компонентов
Закономерности секреции молока
Влияние гормонов на процесс молокообразования
Механизм регуляции молокообразования
Заключение
Литература
Введение
Система молокообразования проявляется в организме периодически после родов и поддерживается в течение всего лактационного периода. Ее исполнительный орган – молочная железа продуцирует сложный биологический секрет – молоко. Основная биологическая роль лактации в эволюционном плане вскармливание потомства.
Функция лактации противоречива по отношению к общей функции организма – самосохранения. В период лактации, особенно при повышении уровня секреции молока, содержащего большое количество питательных веществ, извлекаемых из организма, происходит их истощение, так как включаются мощные нейро-гуморальные механизмы, которые стимулируя секрецию молока, одновременно подавляют механизмы поддерживающие, функцию самосохранения организма.
В период лактации организм животного находится в состоянии нервного напряжения, так как повышается тонус нервной системы, активизируется эндокринная система, усиливаются вегетативные функции организма: увеличивается выделение всех пищеварительных соков, повышается двигательная активность многокамерного желудка и двенадцатиперстной кишки, повышается кровяное давление, усиливается газоэнергетический обмен, увеличивается размер печени.
Под влиянием гормонов в молочной железе формируются основные рабочие структуры – альвеолы с секретирующими элементами и протоковая система. После родов, при отсутствии прогестерона и включении стимулов доения, аденогипофизом выделяются лактогенные гормоны, стимулирующие в первые месяцы лактации более высокий уровень молокообразования. С наступлением новой беременности и началом секреции прогестерона желтым телом процесс молокообразования хотя и не прекращается, но постепенно снижается под влиянием действия гормона прогестерона.
В молочную железу кровью доставляются предшественники молока, из которых в ней заново синтезируются особые сложные белки, жир и молочный сахар – лактоза, а также из плазмы крови отфильтровываются в неизменном состоянии витамины, сывороточный альбумин, лактоглобулин и другие биологически активные и минеральные вещества. Установлено, что оптимальные стимулы доения оказывают более эффективное действие на использование питательных веществ корма для процесса молокообразования. При слабых стимулах доения( машинного) питательные вещества корма в значительной степени используются на отложения в теле животного и не способствуют стимуляции молокообразования, особенно у низкопродуктивных коров. Главным звеном в регуляции лактации является гипоталамо-гипофизарная система, через которую рефлекторно включаются как механизмы рефлекса молокоотдачи, так и механизмы, стимулирующие повышенную секрецию молока после каждого доения. Важным пусковым механизмом для включения этой системы являются стимулы доения. выключение стимулов доения или их ослабление приводит к существенному снижению уровня секреции молока.
Кора больших полушарий двояко действует на функциональную систему лактации: возбужденная кора больших полушарий оказывает тормозящее влияние, невозбужденная кора в состоянии покоя или сна не оказывает тормозящего действия на подкорковые центры и гипоталамус.
Молокообразование сложная многофакторная система, в осуществлении которой принимают участие все функции организма.
Состав молока. Биосинтез его компонентов
Молоко – сложный химический секрет молочных желез, представляет собой полидисперсную среду веществ. Все органические и неорганические компоненты молока, общее количество которых составляет 11-15 %, являются дисперсной фазой и находятся в ионно-молекулярном состоянии и в виде коллоидных и грубодисперсных частиц разной величины. Дисперсной средой молока является вода, количеством 85-89 %. Молоко имеет белый с желтоватым оттенком цвет, сладковатое на вкус и обладающее своеобразным запахом.
В молоке содержится более 100 различных веществ: аминокислоты, жирные кислоты, минеральные вещества, витамины, ферменты, гормоны, углеводы, нейтральные жиры, фосфатиды, стерины, газы и другие вещества. Такие вещества как казеин и лактоза обнаруживаются исключительно в молоке. Водная часть является основным источником воды для новорожденных и средой, форма которой предназначена для самого удобного усвоения молока организмом новорожденного.
Образование молока сложный биологический процесс, который регулируются нервной и эндокринной системами организма. Большинство компонентов молока образуются из приносимых с кровью к молочной железе веществ, некоторые поступают в молочную железу прямо из крови. Поступившие в молочную железу вещества подвергаются в ней сложным химическим превращениям с помощью органоидов секретирующих клеток, они адсорбируются молочной железой и используются для синтеза молочного белка, молочного жира, молочного сахара – лактозы. Количество компонентов, содержащихся в молоке, не постоянно и определяется уровнем кормления и содержания, стадией лактации, уровнем молочной продуктивности, породой, возрастом, временем отела, сезоном года, уровнем нейро-гуморального взаимодействия, наследственностью и так далее.
Белки молока
Белки являются наиболее ценными в питательном отношении. В коровьем молоке их содержится в среднем около 3,2 %. Белки не однородны по своему составу. Основным белком является казеин, или фосфопротеин, который составляет 80 % от общего количества белков молока. По структуре это гетерогенный белок, состоящий из четырех основных фракций: α-казеин, к-казеин, β-казеин и γ-казеин. Кроме фракций казеина в составе молока содержится: α-лактоглобулин, β-лактоглобулин, сывороточный альбумин, иммуно-глобулины.
В целом по содержанию аминокислот белки молока относятся к полноценным белкам (полностью удовлетворяют потребность человека в аминокислотах).
Серосодержащая аминокислота метионин – источник холина и фосфатидов, недостаток их приводит жировому перерождению печени, атрофии эндокринных желез, нарушению передачи нервного возбуждения и к другим расстройствам функций организма.
Цистеин – источник серы и восстановитель, благодаря дисульфидным мостикам молоко при пастеризации приобретает специфический привкус.
Из сыворотки крови в преформированном виде поступают в молоко сывороточный альбумин, γ-казеин, лактоглобулины. Все остальные белки заново синтезируются секретирующими клетками молочной железы. Поэтому из плазмы крови в молочную железу должны непрерывно поступать предшественники для синтеза компонентов молока.
На основании многочисленных исследований установлено, что основными предшественниками синтеза белков являются свободные аминокислоты, поступающие в кровь из пищеварительного тракта при гидролизе белков корма. Белки образуются из аминокислот, синтезируемых микроорганизмами в преджелудках, а также некоторых заменимых аминокислот, синтезируемых в самой молочной железе (вообще клетки молочной железы способны синтезировать все незаменимые аминокислоты, а скорость синтеза β-лактоглобулина и β-казеина зависят от концентрации аминокислот в среде (Schingocthe et al,1967)).
И.К. Медведев, А.К. Швабе (1965) при изучении артерио-венозной разницы в молочной железе установили, что адсорбированный молочной железой азот аминокислот обеспечивает синтез белков молока только на 80 %, а синтез остального количества белков обеспечивается за счет использования белков плазмы крови. И.И. Иванов (1967) на изолированном вымени в условиях перфузии его взвесью отмытых эритроцитов получил молоко почти не отличающееся от натурального молока по составу белков. При перфузии синтез белков молока осуществляется за счет белков самой молочной железы. Для синтеза белков молока молочная железа абсорбирует и комплексные соединения гликопротеинов. Гликопротеины, поступающие в молочную железу, разрушаются, при этом белковая часть используется для синтеза белков, а высвободившиеся углеводы для синтеза лактозы и жира.
В плазме крови значительная часть белков находится в виде комплексных соединений глюцидо-белковых, амино-белковых, минерал-белковых и так далее.
γ-глобулиновая фракция углеводов синтезируется в молочной железе, поступая в общий кровоток, способствует повышению защитных свойств организма в связи с повышением интенсивности обмена веществ и общего его напряжения. α- и γ-глобулиновые фракции – источники как углеводов, так и белков, используемых молочной железой для синтеза лактозы и белков молока. Биосинтез первичных молекул полипептидных цепей – основных белков молока осуществляется на рибосомах эндоплазматического ретикулума из предварительно активизированных лигазами аминоацил-т-РНК-синтетазами свободных аминокислот по общему для всех белков матричному синтезу.
Липиды молока
Липиды молока представлены собственно молочным жиром, фосфолипидами и стероидами. Молочный жир – смесь сложных эфиров трехатомного спирта глицерина и жирных кислот, в нем преобладают олеиновая кислота из ненасыщенных, пальмитиновая, стеариновая, миристиновая – из насыщенных. Наблюдается сравнительно низкое содержание полиненасыщенных жирных кислот: линолевая, линоленовая, арахидоновая, - они регулируют в организме человека липидный, водный и другие обмены веществ, при их недостатке развивается атеросклероз, тромбоз сосудов, сухость кожи и др.
E. Annison et al (1967) установили, что свободные жирные кислоты плазмы крови практически не поглощаются молочной железой. В молочной железе происходит обмен отдельными жирными кислотами между молочной железой и притекающей к ней кровью, поэтому количество жирных кислот не изменяется. Они считали, что жирные кислоты, необходимые для синтеза жира, абсорбируются молочной железой из плазмы крови, а в венозную кровь выводятся не нужные жирные кислоты.
Основными источниками высокомолекулярных жирных кислот молочной железой являются триглицериды плазмы крови, которые находятся в связанном состоянии в виде комплексных соединений липопротеинов.
E.Korn (1963) обнаружил, что в молочной железе содержится фермент липопротеин – липаза, способный гидролизовать липопротеины с высвобождением высокомолекулярных жирных кислот из триглицеридов. Активность фермента резко повышается во время лактации в 3-4 раза, при этом увеличивается его концентрация.
Фракции β-липопротеинов и хиломикронов являются главными поставщиками триглицеридов как источников высокомолекулярных жирных кислот для синтеза молочного жира.
В.И. Никитин (1953) и L.Popjak (1953) установили, что предшественником молочного жира является и глюкоза, из которой в молочной железе образуется вторая часть молекулы жира – глицерин. Предшественниками могут быть и кетокислоты, образующиеся при дезаминировании аминокислотного расщепления белков, и летучие жирные кислоты (уксусная, β-оксимасляная, капроновая), образующиеся в преджелудках в процессе брожения и являющиеся источниками синтеза триглицеридов с низкомолекулярными жирными кислотами.
Углеводы молока
Лактоза – дисахарид, по питательным свойствам не уступает свекловичному сахару, подавляет гнилостные процессы в кишечнике, способствует развитию благоприятной микрофлоры, используется в основном как источник энергии.
Образуется только в молочной железе. Состоит из двух моносахаров – глюкозы и галактозы. Молочная железа активно абсорбирует сахар из плазмы крови. Источником синтеза галактозы является глюкоза, из которой в самой молочной железе под действием комплекса ферментов происходят превращения. При гидролизе гликопротеинов в молочной железе высвобождается галактоза, включающаяся в состав лактозы при ее синтезе. Также предшественниками синтеза лактозы являются низкомолекулярные жирные кислоты (молочная кислота, М. Kleiber, 1954).
Молочная железа активно абсорбирует свободные (в виде моносахаров ) и связанные (в виде гликопротеинов ) углеводы.
Витамины молока
Витамины молока могут быть двоякого происхождения. Одни попадают в организм животного вместе с кормом, другие синтезируются в ЖКТ.
I группа – группа жирорастворимых витаминов:
А (ретинол) и провитамин А (каротин) содержится в зеленом корме
витамины группы D (кальциферолы) образуются при облучении солнцем
Е (токоферол) обладает антиокислительными свойствами
витамины группы F (ненасыщенные жирные кислоты) предупреждают развитие кожных заболеваний и болезней печени.
II группа – группа водорастворимых витаминов:
В1 (тиамин) регулирует углеводный обмен
В2 (рибофлавин) обеспечивает нормальный рост и развитие нервной системы
В12 (кобаламин) участвует в кроветворении
РР (никотиновая кислота) при ее недостатке развивается пеллагра (шершавая кожа)
а также в молоке присутствуют витамины В3, В6, В9, холин, С.
В молоке содержатся все необходимые организму человека витамины, но их количество не может полностью удовлетворить его потребности.
Закономерности секреции молока
Секреция молока в молочной железе в промежутке между доениями, а также на протяжении всего лактационного периода имеет свои закономерности.
Еще в 1924 году C.A. Ragsdale et al. было установлено, что секреция молока постепенно снижается за каждый последующий час после доения и составляет 95% от уровня секреции молока предыдущего часа. L. Bailey (1955) сделал вывод, что секреция молока уменьшается с увеличением интервала между доениями. Авторы признающие постепенное снижение секреции молока в промежутке между доениями, главной причиной такого снижения считают повышение внутривыменного давления по мере накопления молока в вымени – при заполнении системы на 80-85% давление достигает 25 мм.рт.ст. При этом альвеолы растягиваются, железистый эпителий уплощается, и создаются неблагоприятные условия для молокообразования.
В.И. Никитин (1953) установил, что внутривыменное давление в течение 9 часов после доения не изменяется. Это обусловлено тем, что по мере накопления молока в вымени рефлекторно снижается тонус мышечных элементов, при этом увеличивается его емкость, и давление не повышается. Вопрос о закономерности секреции молока в промежутки между доениями недостаточно выяснен. Это объясняется разными методическими подходами, а также трудностями в связи с морфологическими и функциональными особенностями молочной железы. Емкостная система вымени представлена альвеолами, протоками, ходами и цистернами. Емкость ее может значительно изменятся под влиянием тонического рефлекса. Образуемое молоко первоначально накапливается в альвеолах – получить его в первые часы после доения практически невозможно. Из альвеол молоко в протоковую систему и цистерны поступает периодически с неравномерной скоростью и ином составе по сравнению с суммарным (главным образом по содержанию жира). Значит объем остаточного молока и высокое содержание жира могут существенно искажать показатели истинной секреции молока.
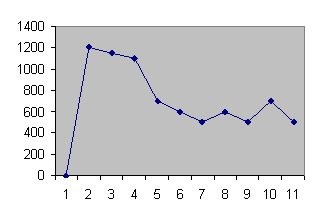
Схема 1. Динамика скорости секреции молока в течение 10 часового промежутка
И.В. Рахманов (1993) провел исследования на лактационных коровах, у которых была проведена полная денервация вымени. В опытный период из вымени выводили только цистернальное молоко, а альвеолярное в течение 10 суток не выводилось – была отключена афферентная импульсация с рецепторов молочных желез. К концу опытного периода секреция молока снизилась на 20%, уменьшилось количество жира, белка и лактозы. Произошло выключение афферентной импульсации с рецепторов молочных желез, а механизмы стимулирующие процессы молокообразования не выключались.
Стимулы доения влияют на повышение выделения лактогенных гормонов в аденогипофизе, которые воздействуют на интенсивность обмена веществ в организме, повышают абсорбирование метаболитов крови молочной железой и секреции молока в первые часы после доения. При выключении стимулов доения секреция молока значительно снижается.
Молочная железа секретирует молоко с неравномерной скоростью. В первые 3-4 часа наблюдается повышение секреции молока, затем она снижается до определенного уровня, в среднем до 580 гр/час и сохраняется на протяжении 10 часов.
Секреция молока состоит из двух фаз:
1 - фаза непрерывной относительно равномерной скорости секреции молока
2 – фаза периодически повышенной секреции молока. Она включается оптимальным стимулом доения в первые 3 часа и наслаивается на фазу непрерывной секреции молока.
Каждая фаза имеет свой, несколько особый, механизм регуляции. Фаза непрерывной секреции регулируется гуморально-гормональным механизмом по принципу моральной связи (Вундер, 1965). Гормон пролактин, поступающий из аденогипофиза в кровь, пройдя по тканям молочной железы, инактивируется. Лактогенные гормоны, пойдя по организму и молочной железе, утрачивают свою активность, и, следовательно, уменьшается их содержание. Такая кровь, поступая в гипоталамус, служит возбудителем хеморецепторов ядер гипоталамуса, в котором образуются нейрогормоны либерины, поступающие в кровь, и по портальной сосудистой системе доставляются в аденогипофиз, в котором стимулируется выделение новой порции лактогенных гормонов.
Эти гормоны, поступая в кровь, стимулируют в организме обмен веществ, в результате чего образуются предшественники молока, используемые молочой железой для синтеза основных компонентов молока, повышают проницаемость мембран секретирующих клеток молочной железы для предшественников молока, активизируют ферментные системы в самих секретирующих клетках, обеспечивающих синтез жира, белка, лактозы и других компонентов.
Фаза повышенной секреции молока включается периодически только стимулами доения. Механизм регуляции этой фазы нейро-рефлекторно-гуморальный. Сущность его состоит в том, что во время акта доения раздражаются рецепторы молочной железы, главным образом механорецепторы сосков, двумя механическими воздействиями: оптимальной силой давления при сжатии сосков и частотой ритма их сжатия. При этих условиях нервные импульсы по афферентным нервным путям поступают в спинной мозг, затем в продолговатый мозг и гипоталамус, стимулируя выделение в кровь повышенной дозы лактогенных гормонов, которые в течение трех часов стимулируют в молочной железе повышенную секрецию молока.
Фазы секреции молока и механизмы их регуляции являются важным физиологическим обоснованием сущности процесса доения, обеспечивающие не только включение механизма рефлекса молокоотдачи и выведение молока, накопление в емкостной системе вымени, но и включение механизмов, стимулирующие последующий процесс повышенной скорости молокообразования. Из этого следует, что раздой коров будет осуществляется в том случае, если после каждого доения более эффективно включается фаза повышенной скорости секреции молока. Если техника доения своим принципом и параметрами не обеспечивает включения фазы повышенной секреции, раздой коров при этих условиях не получается, одновременно происходит и угасание фазы 1, что в конечном итоге приводит к преждевременному прекращению лактации.
Угасание секреции молока на протяжении лактационного периода – закономерное явление. В одних случаях лактационная кривая на протяжении всего лактационного периода поддерживается на более высоком уровне, в других она быстрее угасает, и лактация преждевременно прекращается.
Марченко был проведен опыт на двух группах лактационных коров.
I группа – ручное доение квалифицированной дояркой
II группа – машинное доение
Представленные на схеме 2 и 3 лактационные кривые показывают, что у каждой группы подопытных коров они проходят на разном уровне.
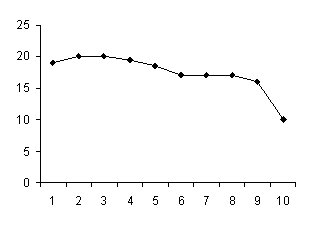 1 – ручное
1 – ручное
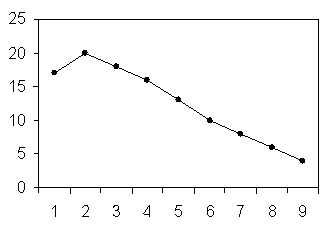 2 - машинное
2 - машинное
Схема 2.Динамика секреции молока при ручном и машинном доении в течение лактационного периода.
Среди лактирующих коров максимальный уровень секреции молока бывает как на первом, так и на третьем месяце лактации. Это обусловлено продолжительностью сервис-периода, то есть временем наступления беременности.
Основным ингибитором лактационной функции организма является гормон прогестерон, который образуется в период беременности в желтом теле яичников.
Гормон прогестерон поступает из желтого тела в кровь, затем в гипоталамус, через который оказывает тормозящее влияние на выделение лактогенных гормонов. При этом снижается метаболизм в целом организме и молочной железе, в частности, что приводит к снижению уровня секреции молока.
Схема 2 показывает, что ручной способ доения является более адекватным и наносит более сильные раздражения на рецепторы сосков за счет за счет силы давления при сжатии сосков и частоты ритма их сжатия (55-80 кПа), по сравнению с машинным способом доения (3-8 кПа).
Сила давления является главным фактором в механическом воздействии на рецепторы сосков, генерирующих эти раздражения в импульсы, которые стимулируют через гипоталамо-гипофизарную систему выделение лактогенных гормонов, а это и обуславливает сильное повышение уровня лактационной кривой на протяжении всего лактационного периода, причем в фазе непрерывной секреции концентрация пролактина в крови выше, чем в фазе повышенной секреции (Протасова Б.И., 1988).
Во второй фазе снижается интенсивность окислительно-восстановительных процессов в молочной железе и в самом организме – активность фермента каталазы при изучении артерио-венозной разницы выше в первой фазе, следовательно, молочная железа активно утилизирует кислород в первой фазе. Во второй фазе снижается чувствительность проводящих нервных путей к стимулам доения.
Более высокий уровень секреции молока в первые месяцы лактации, после родов, обусловлен отсутствием ингибирующего фактора – гормона прогестерона. В последние месяцы лактации, с наступлением беременности и до конца лактационного периода происходит постепенное снижение лактационной кривой под влиянием постепенного поступления в кровь прогестерона из желтого тела яичников. В этот период лактационная кривая поддерживается на определенном уровне под влиянием стимулов доения, которые противодействуют тормозному влиянию гормона.
Молоко, накапливаемое в вымени, на протяжении какого-то промежутка между доениями распределяется в емкостной системе вымени. Молоко, которое накоплено в цистернальной емкости вымени и может быть выведено без акта доения, самотеком, через катетеры, вставленные в сосковые каналы, называется цистернальной фракцией молока. Молоко, содержащееся в альвеолярном отделе вымени, делится на две фракции рефлекторная и остаточная. После выведения цистернальной фракции выводится рефлекорная актом доения, при этом включается рефлекс молокоотдачи. После тщательного выдаивания в альвеолах и протоковой системе содержится некоторое количество молока, называемое остаточной фракцией. Эту фракцию можно вывести с помощью внутривенного введения гормона окситоцина, с последующим его выдаиванием ручным способом.
В цистернальной фракции молока более низкий процент жира, в рефлекторной он значительно повышается и наиболее высокий процент жира в остаточном молоке.
Были проведены исследования влияния ручного и машинного способов доения на эффективность использования питательных веществ потребляемого корма и секрецию молока. Результаты опытов показали, что ручное доение повышает эффективность действия на включение механизмов, стимулирующих лактацию, способствует повышению обмена веществ как в самом организме, так и в молочной железе. При этих условиях питательные вещества корма больше используются молочной железой на процесс молокообразования и меньше на отложения в теле организма.
Влияние гормонов на процесс молокообразования
Под влиянием нейро-гуморального механизма, с наступлением лактации, повышается тонус нервной системы, активизируется эндокринная система, усиливаются вегетативные функции организма, повышается интенсивность абсорбирования метаболитов молочной железой, активизируются ферментативные системы молочной железы, принимающие участие в синтезе составных частей молока.
В регуляции лактации принимает участие одновременно целый комплекс гормонов, почти всей гаммы желез внутренней секреции.
Роль пролактина
Гормон образуется в передней доле гипофиза и представляет собой белок с одиночной пептидной цепью. Его образование находится в обратной зависимости от содержания в гипоталамусе дофалина и образования надпочечниками гормона кортизола. Стимулы доения оказывают последействие на повышение интенсивности выведения пролактина из аденогипофиза, что обуславливает повышение уровня секреции молока в первые часы после доения.
Пролактин обладает полифункциональным действием на процессы в лактирующем организме: тормозит в гипофизе выделение фолликулостимулирующих гормонов, влияет на активность железистого эпителия в молочной железе, ограничивает поступление глюкозы из крови в ткани, увеличивает концентрацию свободных жирных кислот, повышает синтез ДНК, активность фермента РНК-азы, синтез РНК, оказывает непосредственное влияние на секретирующие клетки молочных желез, активизирует фермент липопротеинлипазу.
Пролактин обнаруживается в связи с митохондриями и микосомами секретирующих клеток – происходит изменение активности ферментов, связывание с белками, обуславливающими секретирующие активность молочной железы.
Опыты показали, что после введения пролактина козам в 1 и 2 периодах лактации повысилось содержание углеводов в артериальной крови. Введение пролактина во второй фазе оказало более эффективное действие – на фоне снижения гормонального статуса в крови, процессы в молочной железе и реакции лактирующего организма оказались более интенсивными. Пролактин повышает чувствительность секреторного эпителия молочной железы к действию этого гормона.
Содержание пролактина в крови в первом периоде выше (около 95-100 нг/мл), чем во втором (около 30нг/мл) (Б.И. Протасова, 1988).
Можно считать, что пролактин стимулирует абсорбирование молочной железой четырех фракций гликопротеидов – альбуминовой, α1-α2-β-глобулиновых. Поступающие фракции гликопротеинов в молочной железе, гидролизуясь, служат источниками белковых структур, используемых для синтеза белков молока, а также углеводов в виде глюкозы и галактозы для синтеза лактозы и молочного жира.
Фракция α-липопротеинов синтезируется молочной железой, а β-липопротеинов используется ею как источник высокомолекулярных жирных кислот для синтеза молочного жира, при этом фермент липопротеинлипаза осуществляет гидролиз липопротеинов на белковые структуры, триглицериды с высвобождением высокомолекулярных жирных кислот.
Пролактин оказывает стимулирующее действие на процессы в самом организме и биохимические в молочной железе: повышает проницаемость мембран секретирующих клеток (Яковлев и др., 1973), активизирует липопротеинлипазу и лактазную синтетазу, обмен углеводов, фракции глико- и липопротеинов, их абсорбирование молочной железой.
Роль соматотропного гормона
Гормон соматотропин полипептид, обладающий видовой специфичностью. Он повышает в клетках синтез рибосомальной и транспортной РНК, регулирует жировой обмен, стимулирует выделение гормона глюкагона в поджелудочной железе, влияет на утилизацию глюкозы жировыми тканями, где она используется как энергетический материал и источник для синтеза жира, повышает концентрацию летучих жирных кислот в рубце (особенно уксусной), увеличивает содержание в плазме крови глюкозы, фосфатидов и нейтрального жира (Willimsonet, 1963), стимулирует образование в печени и почках соматомедина, который влияет на белковый обмен, ускоряет рост и развитие молочных желез (Li, 1969), усиливает превращение веществ корма в молоко, то есть повышает коэффициент полезного действия корма (Mashin, 1973), то есть соматотропин влияет на различные стороны белкового, жирового и углеводного обмена, что приводит к усиленному образованию молока – к росту молокообразования.
Ф.Ю. Рышка (1988) выявили при исследовании аденогипофиза, что гормон обладает двойным действием соматотропным и лактогенным, и назвали лактосоматотропин.
Соматотропин активизирует и тормозит ферментные системы, изменяет проницаемость клеточных мембран, активизирует генетические процессы в ядре, понижает выделение с мочой солей K, Na, Mg, Cl, P и повышает выделение солей Са. Он выделяется больше ночью, чем днем, до 90 % его расщепляется в печени, а остальное в почках и других паренхиматозных органах, оказывает более мощное действие на метаболические процессы в лактирующем организме и непосредственно на функцию лактации по сравнению с действием пролактина.
Соматотропин, с одной стороны, повышает содержание фракций гликопротеинов (альбуминовая, α1-α2-β-глобулиновые) и их абсорбцию молочной железой, а с другой стороны, стимулирует образование γ-глобулиновой фракции в молочной железе, которая, поступая в кровяное русло, способствует повышению иммунных свойств организма.
В первом периоде лактации, даже при повышении гормонального статуса, соматотропный гормон оказывает стимулирующее действие на липопротеиновый обмен, во втором - еще большее стимулирующее действие на такой обмен и активность их абсорбирования молочной железой фракции β-липопротеинов, одновременно повышает синтез в молочной железе и выведение в венозную кровь фракции α-липопротеинов.
Таким образом, соматотропный гормон является одним из важных лактогенных гормонов, участвующих в лактации.
Роль тироксина
Тироксин – основной гормон, образующийся в щитовидной железе, который входит в комплексное соединение – тиреоглобулин, накапливаемый в полостях фолликулов щитовидной железы. Под действием тиреотропного гормона передней доли гипофиза, поступающего с кровью в щитовидную железу, активизируются ферменты пептидазы и протеазы, которые гидролизуют тиреоглобулин и освобождают из него гормоны тироксин и трийодтиронин. Эти гормоны из фолликулярных полостей абсорбируются с помощью микроворсинок на апикальной стороне эпителиальных клеток и выводятся в кровь через базальную сторону клеток.
Поступившие в кровь гормоны связываются с белками альбуминами и α-глобулинами плазмы крови, образуя связанный с белком крови йод (СБЙ). В составе СБЙ тироксин составляет 80-90 % его содержания в крови, после его поступления в клетки тканей он превращается трийодтиронин, который в 5-6 раз активнее тироксина.
Местом приложения гормонов щитовидной железы являются митохондрии клеток, в которых они стимулируют процессы окисления за счет повышения реактивной способности дисульфидных групп, повышают проницаемость мембран митохондрий и входящих в них субстратов цикла трикарбоновых кислот, усиливают расход АТФ, вызывают разобщение окислительного фосфорилирования, при этом происходит окисление фосфогена и молочной кислоты и образуется тепло.
Тироксин обладает свойством полифункционального действия: метаболическое – стимулирует интенсивность белкового, жирового и углеводного обменов, оказывает влияние на водно-солевой; морфогенетическое – регулирует формообразовательный процесс, воздействует на генетические структуры; всасывающее – изменяет проницаемость клеточных мембран, усиливает или подавляет процесс всасывания и реабсорбции.
В период лактации наблюдается гиперфункция щитовидной железы, особенно у высокопродутивных животных, что приводит к напряжению ЦНС.
Действие гормона тироксина на содержание различных гликопотеиновых фракций близко к действию на их содержание гормона соматотропина, то есть происходит повышение содержания альбуминов, α1-α2-β-глобулинов, в то время как γ-глобулины обнаруживаются в венозной крови, оттекающей от молочной железы. Также после введения тироксина повышается содержание в артериальной крови фракций α-липопротеинов, β-липопротеинов и хиломикронов, они играют важную роль в лактации как источники высокомолекулярных жирных кислот и аминокислот.
Роль инсулина
Гормон инсулин образуется β-клетками островков Лангерганса поджелудочной железы. Стимулирование синтеза инсулина происходит в основном при повышенной концентрации глюкозы в крови. Он стимулирует синтез ферментов гексокиназы и глюкокиназы, которые фосфорилирует глюкозу, в результате образуется глюкозо-6-фосфат, поступающий в печень и используется для синтеза гликогена. Инсулин стимулирует синтез информационной РНК, активизирует цикл Кребса, усиливает утилизацию глюкозы с образованием жира.
Инсулин угнетает секрецию молока и лактозу, но повышает секрецию молочного жира и казеина, оказывает существенное влияние на метаболизм в организме.
В результате опытов было установлено, что при введении повышенной дозы инсулина не оказало влияния на секрецию молока. Среднесуточный удой практически сохранился на исходном уровне, однако, повысилась секреция молочного жира и молочного белка. Это объясняется тем, что повышенная доза инсулина по принципу обратной связи оказывает стимулирующее действие на увеличение выделения соматотропных гормонов, которые стимулируют синтез белков ( Балаболкин, 1969).
Роль эстрогенных гормонов
Половые гормоны по химической природе относятся к стероидам. Основным источником их образования является холестерин. К половым гормонам самки относятся эстрадиол и его производные – эстрон и эстриол, образующиеся в яичниках, прогестерон и релаксан в желтом теле яичников, лактотропин и хориогонин – в плаценте.
Эстрогены по своему действию являются полифункциональными гормонами. Они стимулируют рост и развитие половой системы самки, повышают синтез белков в организме и окисление жиров и углеводов. Одновременно снижается уровень холестерина и β-липопротеинов.
Эстрогены, циркулирующие в крови, находятся в свободном и связанном с белками состоянии. В печени они инактивируются и разрушаются. Оказывают тормозящее влияние на секрецию фолликулостимулирующего гормона и стимулируют секрецию лютенизирующего и лактотропного гормонов. Во время беременности повышенная доза эстрогенов тормозит секрецию этих гормонов. В молочной железе эстрогены влияют на рост и формирование протоковой системы. При длительном воздействии они оказывают тормозящее действие на гипоталамус и аденогипофиз, затормаживает выделение лактогенных гормонов – пролактина, соматотропина и тиреотропина. Таким образом являются ингибиторами функции лактации (Hutton,1958).
Стероидные гормоны имеют относительно невысокий молекулярный вес, поэтому проникают через мембрану клитки в цитоплазму, а затем и в ядро и там реализуют свой эффект.
Механизмы регуляции молокообразования
Нервная регуляция
В регуляции непосредственной секреторной функции молочной железы главную роль играют гормоны желез внутренней секреции. Это обусловлено тем, что молочная железа иннервируется только симпатическими нервами, которые обеспечивают афферентную связь молочной железы с ЦНС за счет разных рецепторов в молочной железе и эфферентную – регулирующую сосудистую и гладкомышечную протоковые системы молочной железы. Но эти нервы не обладают стимулирующим свойством секреторной функции молочной железы. В молочной железе отсутствует парасимпатическая иннервация, обладающая свойством стимуляции секреторного процесса (Богданов, 1957).
Молочная железа в процессе эволюции является более поздним образованием – она возникла, когда ЦНС млекопитающих достигла высокой степенью развития и дифференциации. Отсутствие парасимпатической иннервации явилось биологической необходимостью выживания вида млекопитающих. На протяжении многих веков, находясь в дикой природе, эти животные подвергались стрессовым воздействиям различных природных явлений, а также нападению хищников.
В этих условиях самка пытается вместе с потомством скрыться от хищника, находясь в состоянии стресса. В таких случаях, если бы молочные железы иннервировались парасимпатическими нервами, регулирующие ее секреторную функцию, то это вызвало бы полное торможение секреции молока. Одновременно потомок, убегая вместе с матерью, теряет много энергии, ему требуется срочное восстановление ее за счет приема молока, которое в молочной железе отсутствует. В таких условиях произошло бы вымирание вида. Потому в процессе эволюции сформировались млекопитающие животные, у которых молочная железа оказалась отключенной от парасимпатической иннервации, а регуляция ее секретирующей функции стала осуществляться гормональным механизмом ЖВС через гипоталамус. Этот механизм включается и поддерживается автоматически, по замкнутому кругу, по принципу обратной связи. При выделяемые в кровь лактогенные гормоны оказывают непрерывное стимулирующее действие как на вегетативные функции организма, так и на секретирующую функцию молочной железы более продолжительное время по сравнению с кратковременным действием нервных импульсов, что обеспечивает сохранение данного вида животных даже в условиях стресса.
Гуморальная регуляция
Гипоталамо-гипофизарная система представлена двумя взаимосвязанными структурами – гипоталамусом и гипофизом посредством портальной сосудистой системы.
Гипоталамическая область связана афферентными путями со спинным, продолговатым и средним мозгом, отдельными полями коры больших полушарий и другими мозговыми структурами, через которые теоретически допускается возможность получения информации гипоталамусом от любой части мозга (Ахилла, 1981).
Нейроны гипоталамической зоны передних ядер подбугорья являются местом трансформации афферентных нервных стимулов в гормональные факторы, которые обладают высокой специфической активностью. К ним относятся рилизинг-факторы: либерины и статины, влияющие на секреторную функцию аденогипофиза. Либерины стимулируют секрецию гормонов аденогипофиза, а статины тормозят.
Либерины: пролактолиберин, соматолиберин, тиреолиберин и др.
Статины: соматостатин, пролактостатин, меланостатин.
Рилизинг-гормоны выделяются в кровь и по сосудисто-портальной системе поступают в передней и средней доли гипофиза. Поступившие в переднюю либерины повышают проницаемость клеточных мембран секретирующих клеток аденогипофиза посредством адренатциклазной системы, при этом в секретирующие клетки проникают катионы Са, которые активируют освобождение гормонов из секретирующих клеток аденогипофиза.
Стимулирующее действие на образование и выделение рилизинг-гормонов либеринов в гипоталамусе оказывает медиатор серотонин, которые образуются из аминокислоты триптофана. Он тормозит выделение адреналина, норадреналина, мелатонина и дофамина, тормозящих образование либеринов и ингибирующих действие прогестерона и эстрогенов в гипоталамусе.
Гипоталамо-гипофизарная система регулируется по принципу обратной связи. Ядра гипоталамуса имеют рецепторы, которые чувствительны к изменению концентрации гормонов в крови. При повышении концентрации наступает торможение выделения этих гормонов и наоборот.
В гипоталамусе образуются два гормона – окситоцин и вазопрессин, которые поступают в заднюю долю гипофиза, откуда (под влиянием стимулов доения) выводятся в кровь. Окситоцин поступает в молочную железу, действует на миоэпителий альвеол и вызывает молокоотдачу, а вазопрессин, обладая антидиуретическим свойством, влияет на процесс мочеобразования, повышает реабсорбцию воды в канальцах почек, при этом уменьшается выделение воды с мочой. В организме после доения задерживается вода, которая необходима молочной железе для молокообразования.
Роль стимулов доения на процесс молокообразования
Под влиянием стимулов доения повышается выделение лактогенных гормонов из аденогипофиза, повышается деятельность вегетативных функций лактирующего организма, в частности газообмен, двигательная активность желудка и двенадцатиперстной кишки, повышается кровяное давление, усиливается и учащается дыхание. Для поддержания секреции молока требуется периодическое сосание или доение. Прекращение подобных раздражений молочной железы приостанавливает отделение молока вне зависимости от периода лактации. Многократное доение в течение дня значительно повышает количество молока в сутки.
Роль коры больших полушарий в регуляции лактации
По данным Протасова Б.И. (1988), кора больших полушарий имеет прямой контакт с латеральным, маммилярным, вентромедиальным и паравентрикулярным ядрами гипоталамуса, в которых образуются рилизинг-гормоны либерины. В случае удаления влияния коры больших полушарий на нижележащие отделы ЦНС происходит повышение секреции молока.
Высокопродуктивные коровы находятся словно полусонном состоянии – они больше лежат и спят. В данном случае кора больших полушарий находится в состоянии торможения и не оказывает тормозного влияния на ядра гипоталамуса и на его вегетативные центры, что способствует активизации секретирующего процесса в молочной железе. У малопродуктивных коров кора находится в состоянии возбуждения, и оказывает тормозящее влияние на ядра гипоталамуса.
Ярким примером влияния возбуждения коры больших полушарий на торможение синтеза молока является случай, описанный И.Д. Рихтер (1939), когда перед глазами отелившейся коровы зарезали рожденного ею теленка. При этом корова сильно металась, мычала, а затем у нее полностью прекратилась лактация.
Учитывая эту зависимость очень важно правильно оценить и подобрать технологические процессы на молочной ферме. Прежде всего необходимо учитывать общий биологический «закон» - «борьбу индивидуумов за территориальную неприкосновенность». Известно, что коровы при привязном содержании, приученные к своему стойлу, не допускают к этому месту других животных и могут спокойно лежать, спать, поедать корм. Процесс доения доставляет корове приятное чувство. Она закрывает глаза и как бы временно отключает себя от внешних воздействий. Это способствует более эффективному проявлению рефлекса молокоотдачи и активному включению механизмов, стимулирующих последующий процесс молокообразования под влиянием стимулов доения.
При технологии группового безпривязного, боксового содержания, группового кормления и доения, когда животные свободно располагаются на ограниченной площади, при этом происходит разделение на сильных и слабых, они постоянно находятся в определенном нервном напряжении.
Вывод заключается в том, что на молочных фермах система технологического процесса должна быть такой, чтобы на животных действовало меньше всяких внешних и внутренних раздражителей, возбуждающих кору больших полушарий.
В ночной период многие внешние раздражители исключаются, животные находятся в состоянии покоя или сна, при этих условиях функционируют подкорковые центры и гипоталамус, что и обуславливает повышенную активность секреторного процесса в молочной железе, животные больше времени затрачивают на жвачку – скорость потребления корма понижается, а усвояемость повышается.
Молокоотдача
Молокоотдача – это активное изгнание молока, накопленного в емкостной системе вымени, при включении рефлекса молокоотдачи под влиянием акта сосания или доения.
Включается рефлекс нейрогуморальным механизмом и при раздражении рецепторов сосков. В молочной железе имеются механорецепторы, прессорецепторы, тактильные рецепторы, барорецепторы, хеморецепторы и терморецепторы (Грачев, 1952).
При доении механические и термические воздействия на рецепторы сосков трансформируются в нервные импульсы, поступающие по афферентным нервам в спинной мозг, из которого обратно по эфферентным волокнам нервные импульсы поступают в молочную железу и вызывают расслабление гладкой мускулатуры протоковой системы и сфинктеров сосков. Это первая нейрогуморальная фаза рефлекса молокоотдачи (она длится 5-15 секунд). Вторая фаза: поступающие нервные импульсы в спинной мозг одновременно передаются в продолговатый мозг, а затем по другим нейронам поступают в гипоталамус.
Латентный период (период от начала раздражения рецепторов сосков до начала поступления в молочную железу гормона окситоцина и сокращения миоэпителия альвеол) имеет оптимальное время 25 секунд.
При ручном доении он колеблется в пределах от 25 до 43 секунд, а при машинном 50-132 секунды. Следовательно, одна из главных причин заболевания вымени коров маститом является «холостое» доение – при большой продолжительности латентного периода и при передержки доения доильным аппаратом.
Заключение
Несмотря на то, что молоко образуется молочной железой, в формировании его как специфического секрета участвует весь организм. Лактация есть функция всего организма. Образование молока эпителиальными клетками в молочной железе обеспечивается при участии многих систем организма: пищеварительной, дыхательной, половой, выделительной, эндокринной, нервной, газоэнергетического обмена и кровеносной. Единство всех систем организма обеспечивается нервной системой с ее высшим отделом - головным мозгом.
На основе оценки типов нервной деятельности по Павлову можно судить о лактогенной функции молочной железы. Коровы сильного уравновешенного подвижного типа НС имеют более высокие адаптационные, защитные и компенсационные способности. У них формируются мощные системы условно-безусловного рефлексов молокоотдачи, способные противостоять тормозным воздействиям окружающей среды, которые обеспечивают высокое постоянство удоя в течение лактации, меньшие суточные колебания удоя, более рационального распределения энергии корма. Для коров слабого типа НС характерна слабость возбудимость и низкая подвижность нервных процессов. Раздой проходит у них медленнее, ярко выражены суточные колебания удоя, поскольку любые тормозящие воздействия снижают удой, а восстановление его проходит медленно.
К началу 70-х годов в науке о лактации уже существовали многочисленные доказательства важной роли аденогипофиза и прежде всего пролактина – основного лактогенного гормона в регуляции лактационной функции организма, включая маммогенез, лактогенез и лактопоэз. При доении происходит резкое увеличение уровня пролактина в крови, он стимулирует абсорбирование молочной железой углеводов и белков. Похожее действие оказывают на молочную железу соматотропин и тироксин. В то же время гормон инсулин угнетает секрецию молока, а эстрогенные гормоны затормаживают выделение лактогенных гормонов.
Млекопитающие приобрели огромное преимущество в процессе эволюции по сравнению с другими видами. Они получили возможность вскармливать детенышей молоком – очень ценным и полезным продуктом, особенно для растущего организма – в любое время, независимо от условий внешней среды, стрессовых воздействий, нехватки пищи. В молоке содержатся все необходимые вещества, которые требуются для полноценного развития организма, потому что многие минералы, витамины, аминокислоты и др. требуются в первые месяцы жизни малыша, и он их получает. Вследствие этого млекопитающие заняли высшую ветвь древа эволюции.
Литература
1.«Биологические основы высокой продуктивности». Боровск, 1990.-с.64-72
2.Галанцев В.П., Гуляева Е.П. «Эволюция лактации». Л.: Наука, Ленингр. отд., 1987.-248 с.
3.Грачев И.И. «Клеточные механизмы регуляции секреции молока». М.: Элиста, 1995.-95 с.
4.Грачев И.И. «Современные достижения физиологии и биохимии лактации». Л.: Наука, 1991.-123 с.
5.Кокорина Э.П. «Условные рефлексы и продуктивность животных». М.: Агропромиздат, 1996.-196 с.
6.Кобянов Л.С. ст. «Уровень инсулина у лактирующих коз». Труды Т29 «Физиология пищеварения и лактации». Алма-Ата, 1986.- 171 с.
7.Макар З.Н., Черепанов Г.Г. ст. «Взаимосвязь органного кровотока активности транспорта в секретирующие клетки молочной железы и образование компонентов молока у коров». Рос. физиол. журн. им. Сеченова Т89 8, Л.: Наука
8.Погодюк П.З. ст. «Повышение молокообразования при введении коровам гормона роста». Журн.: Вестник с/х науки №7, 1990
9.Протасов Б.И. «Исследование гипоталамической регуляции секреции молока у жвачных». Автореф. дисс. д-ра биол. наук. Боровск, 1987
10.Тараненко А.Г. «Физиологические основы повышения молочной продуктивности». М.: «Россельхозиздат», 1997.- 130 с.
11.Федосимов В.А., Кокорина Э.П. ст. «Исследования по физиологии и биохимии лактации коров». Журн. Зоотехния, 8.2000
© 2010 Интернет База Рефератов