
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Реферат: Передача и кодирование сигнала в сетчатке глаза
Реферат: Передача и кодирование сигнала в сетчатке глаза
Передача и кодирование сигнала в сетчатке глаза
Содержание
Передача и кодирование сигнала в сетчатке глаза
Глаз
Анатомия проводящих путей зрительного анализатора
Палочки и колбочки
Организация и морфология фоторецепторов
Электрические сигналы в ответ на свет в фоторецепторах позвоночных
Слуховая кора
Локализация звука
Выводы
Литература:
Передача и кодирование сигнала в сетчатке глаза
Путь нервных импульсов, возбуждаемых светом и позволяющих нам воспринимать окружающие нас объекты, фон, движение, тени и цвета, начинается на сетчатке. Свет воспринимается специальными фоторецепторами, колбочками и палочками, содержащими зрительные пигменты. Палочки высокочувствительны и способны уловить даже один квант света. Цветовое дневное зрение осуществляется колбочками. Поглощение света зрительным пигментом фоторецептора активирует G-белок, запускающий каскад биохимических реакций. В результате происходит закрытие управляемого нуклеотидами ионного канала, расположенного на мембране фоторецепторной клетки, что приводит к ее гиперполяризации. Таким образом, при действии света происходит уменьшение высвобождения медиатора в синапсах между фоторецепторами и биполярными клетками, а также между фоторецепторами и горизонтальными клетками. Сигналы от фоторецепторов достигают в конечном итоге ганглиозных клеток, аксоны которых проходят в составе зрительного нерва и являются единственными нервными волокнами, несущими зрительную информацию в мозг.
Соединения между рецепторами и ганглиозными клетками обеспечиваются биполярными, горизонтальными и амакриновыми клетками. Подобно палочкам и колбочками, биполярные и горизонтальные клетки генерируют сигналы в виде локальных ответов, а не потенциалов действия. Кодирование зрительных сигналов в сетчатке и на последующих уровнях зрительного анализатора можно лучше всего изучить при помощи анализа нейронных механизмов функционирования рецептивных полей, являющихся элементарными блоками, из которых строится система восприятия зрительных образов. Рецептивным полем нейрона зрительного анализатора называется определенная зона сетчатки, при освещении которой изменяется (увеличивается или уменьшается) активность этого нейрона. Рецептивным полем ганглиозной клетки сетчатки является небольшая округлая область сетчатки. Потенциалы действия возникают в ганглиозных "оп"-клетках, воспринимающих маленькие пятна света, расположенные в центре темного поля, и в ганглиозных "off''-клетках, воспринимающих темные пятна на освещенном фоне. Эти две группы ганглиозных клеток имеют большое значение в работе сетчатки. Называемые мелкоклеточными (parvocellular. Ρ) и крупноклеточными (magnocellular, M), они имеют разные размеры, расположение, связи и физиологические ответы. Более мелкие Ρ клетки способны к высокоточной пространственной дискриминации и способны различать различные цвета. Большие по размеру M клетки способны воспринимать подвижные объекты, а также чувствительны к малейшим изменениям контрастности. Эти свойства M и Ρ групп клеток передаются по нервным связям на дальнейшие уровни зрительного анализатора, вплоть до уровня сознания.
Глаз
Глаз - автономный аванпост головного мозга. Он собирает информацию, анализирует и передает ее для дальнейшего анализа в высшие центры по строго определенному пути (зрительному нерву). Первым шагом здесь является получение перевернутого изображения окружающего мира на сетчатке. Неотъемлемыми условиями четкого зрения являются: (1) фокусирование изображения на сетчатке путем изменения толщины хрусталика (аккомодация); (2) регуляция зрачком количества света, падающего на сетчатку; (3) конвергенция информации от обоих глаз для того, чтобы изображения на сетчатках обоих глаз располагались в соответствующих зонах. Наше зрение сильно зависит от области зрительного поля, на которое проецируется изображение. Мы способны читать мелкий текст, расположенный прямо перед глазами и проецирующийся в центр сетчатки, но не куда-то в периферическую зону. Падение остроты зрения возникает из-за различных способов анализа получаемой зрительной информации в разных зонах, а не из-за нерезкого изображения или оптических аномалий вне центральной зоны. Сначала мы познакомимся с принципиальными анатомическими особенностями пути зрительного анализатора, а затем шаг за шагом рассмотрим, каким образом возникает сигнал на сетчатке при улавливании света зрительными пигментами и передается затем далее по зрительному пути.
Анатомия проводящих путей зрительного анализатора
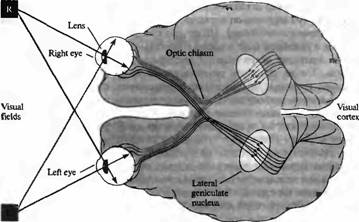
Путь зрительного анализатора, начинающийся от глаза и идущий до коры головного мозга, показан на рис.1, где изображены основные ключевые пункты зрительной системы. Волокна зрительного нерва начинаются от ганглиозных клеток сетчатки и заканчиваются в слое клеток таламуса, в области латерального коленчатого тела, где расположены вторые нейроны цепочки зрительного тракта. Аксоны нейронов коленчатого тела, в свою очередь, проходят через зрительную лучистость в кору головного мозга. С этого этапа распространение зрительной информации происходит еще более сложным путем, проследить который пока не представляется возможным
На рис.1 показано, как волокна, идущие от каждой сетчатки, разделяются на два пучка в области зрительного перекреста, откуда они идут в латеральное коленчатое ядро и зрительную кору обоих полушарий. Правые стороны сетчатки обоих глаз посылают волокна в правое полушарие. Из-за того, что изображение переворачивается, проходя через зрачок, получается, что на правую сторону каждой сетчатки проецируется изображения окружающего мира с левой стороны от животного. Таким образом, каждое полушарие "видит" зрительное поле противоположной стороны. Соответственно, люди с повреждением левого полушария из-за травмы или заболевания становятся "слепыми" на правое.
Рис.1. Путь зрительного анализатора. Правая сторона сетчатки каждого глаза, показанная серым цветом, посылает сигналы к правому латеральному коленчатому телу. Таким образом, зрительный центр правого полушария получает информацию исключительно о левой части зрительного поля.
Таковыми являются горизонтальные и амакрииовые клетки. Только амакриновые и ганглиозные клетки способны отвечать потенциалом действия и передавать его на другие клетки. Фоторецепторы, горизонтальные и биполярные клетки отвечают только локальным ответом переменной амплитуды. Внутри каждого из этих основных классов имеются подгруппы, отличающиеся друг от друга по строению и функциям, мюллеровские клетки являются глиальными клетками сетчатки.
Палочки и колбочки
Фоторецепторы являются теми образованиями, которые позволяют нам видеть окружающий мир, а также определяют диапазон, в пределах которого мы способны воспринимать его. Некоторые змеи имеют специализированные рецепторы для восприятия инфракрасного излучения; муравьи и пчелы способны для навигации использовать свойства поляризованного света неба. Наши фоторецепторы не способны ни к первому, ни ко второму. Кошки, не имеющие соответствующих рецепторов, не способны воспринимать цвета. Они "страдают" цветовой слепотой, воспринимая все цвета подобно тому, как мы видим в сумерках (когда "все кошки серы"). Чувствительность наших палочек в темноте такова, что один квант света способен вызвать поддающийся измерению сигнал. Однако около 7 палочек должно быть активировано, чтобы мы действительно были способны воспринять свет.
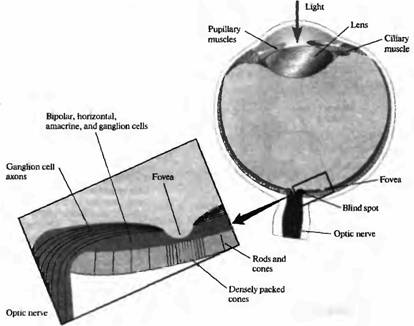
Рис. 3. Путь света в полости глаза и клеточное строение сетчатки (поперечный срез глаза). Свет должен пройти через хрусталик и слои клеток сетчатки, чтобы достигнуть фоторецепторов: палочек и колбочек. Центральная ямка (fovea centralis) является специализированной зоной, содержащей только густо расположенные тонкие колбочки, которые способны различать объекты с высоким разрешением. В этом месте слои клеток, расположенные более поверхностно, менее выражены, что позволяет свету непосредственно падать на фоторецепторы. В месте, где зрительный нерв выходит из глаза, отсутствуют фоторецепторы, и потому это место называется "слепым пятном".
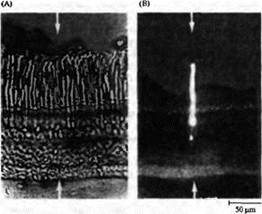
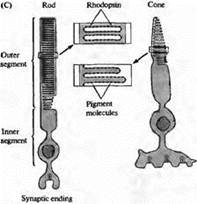
Рис.4. Фоторецепторы сетчатки. (А и В) Палочка сетчатки жабы с введенным в нее флуоресцентным красителем, Lucifer yellow, при обычном (А) и ультрафиолетовом (В) облучении. Стрелками отмечены одинаковые места в сетчатке. (С) Строение палочки и колбочки. В палочке пигмент родопсин (черные точки) встроен в мембраны, расположенные в виде дисков, не связанных с наружной мембраной клетки. В колбочке зрительные пигменты располагаются в складках наружной клеточной мембраны. Наружный сегмент связан с внутренним посредством узкой перемычки. В темноте происходит постоянное высвобождение медиатора нервными окончаниями.
При помощи различных подтипов колбочек мы способны воспринимать мельчайшие оттенки и изменения в контрастности и цвете в яркий солнечный день, когда интенсивность света в 100 миллионов раз сильнее, чем ночью.
Организация и морфология фоторецепторов
Палочки и колбочки образуют плотно организованный слой фоторецепторов, расположенный непосредственно над слоем пигментного эпителия (рис.3), на максимальном удалении от роговицы и проникающего в глаз света. За исключением небольшой зоны в области центральной ямки, свет должен пройти через несколько плотных слоев клеток и волокон, чтобы достигнуть наружного сегмента фоторецептора, где и осуществляется поглощение фотонов и фоторецепция. Как Гельмголъц писал в 1867открыт синапс и вообще появилась клеточная доктрина*).
Исследования центральной ямки показали, что в этой зоне колбочки очень плотно упакованы, их плотность составляет 200000 на мм2, а палочки отсутствуют. Более того, в области ямки колбочки более узкие, чем в периферических зонах сетчатки. Так как ямка не содержит палочек, ночью она представляет собой "слепое пятно". Существует и отдельное "слепое пятно", которое соответствует зоне на сетчатке, где зрительный нерв покидает глаз; в зоне этого пятна отсутствуют любые фоторецепторы.
На рис.4 показаны три важные особенности строения фоторецепторов: (1) наружный сегмент, где происходит поглощение света зрительными пигментами; (2) внутренний сегмент, где расположены ядро, ионные насосы, трансмембранные переносчики, рибосомы, митохондрии и эндоплазматический ретикулум; и (3) синаптическая терминаль, где происходит высвобождение глутамата на клетки второго порядка, чувствительные к глутаматному сигналу. Зоны секреции в синаптической терминали имеют строго определенное расположение в зависимости от расположения одной или нескольких "ленточных" структур, вдоль которых ориентированы везикулы с медиатором.
Хотя сама по себе "клеточная доктрина", или "клеточная теория", была опубликована Маттиасом Шлейде ном и Теодором Шванном в 1839 году ("Микроскопические исследования о соответствии в структуре и росте животных и растений"), она содержала несколько ошибочных утверждений (например, о новообразовании клеток организма из первичного неклеточного вещества), и получила известность и распространение значительно позже, после уточнения ее Рудольфом Биржевым в 1859 году
Электрические сигналы в ответ на свет в фоторецепторах позвоночных
Рецепторы в основном отвечают на соответствующий стимул путем локальной деполяризации переменной амплитуды, которая может вызвать в дальнейшем потенциал действия. И хотя у большинства беспозвоночных фоторецепторы воспринимают свет подобным образом (рис.5А), восприятие света рецепторными клетками у большинства позвоночных происходит по-другому. На рис.5В показаны сигналы, отведенные от палочки черепахи при помощи внутриклеточного электрода. В темноте (во время отдыха) фоторецепторы деполяризуются постоянным входящим током в области наружного сегмента ("темновой ток"). Свет приводит к гиперполяризации, уменьшая величину входящего тока. В следующих абзацах будет рассказано, каким образом свет поглощается фоторецепторами и какие механизмы вызывают электрические сигналы, показанные на рис. 5.
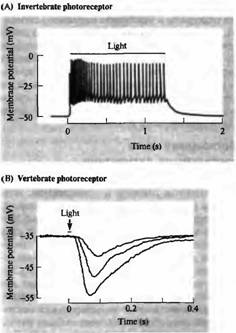
Рис. 5. Сигналы от фоторецепторов. (А) Фоторецепторы беспозвоночных (мечехвост) отвечают на свет деполяризацией, вызывающей серию разрядов. Такие ответы являются типичными для сенсорных клеток, чувствительных к различным стимулам: прикосновению, давлению или растяжению (глава 17). (В) Фоторецепторы позвоночных (черепахи) отвечают на свет гиперполяризацией, уровень которой зависит от интенсивности вспышки.
Слуховая кора
Слуховой вход, осуществляемый как через дорзальные, так и вентральные кохлеарные ядра, достигает слуховой коры. Первичная слуховая кора (А,) расположена в верхнем отделе височной доли и соответствует полям 41 и 42 по Бродману. У кошек А, удобно расположена на боковой поверхности мозга, поэтому большинство комбинированных анатомо-физиологических исследований производится на этом виде. Микроэлектродные исследования показали, что А, также имеет колончатую организацию, при которой все клетки, расположенные вдоль вертикального пути, имеют одну и ту же оптимальную частоту. Фактически A1 организована в виде изочастотных полос или пластин коры, идущих перпендикулярно к тонотопической оси.
Слуховая кора у обезьян содержит три полных кохлеотопических карты с параллельными проекциями из ядер медиального коленчатого тела на все эти области. Самая задняя из них соответствует первичной слуховой коре, как показано на основе ее гистологических свойств. Эта центральная основа окружена вторичными слуховыми областями, которые соединены не только с первичной корой, но также и с подразделами ядер медиального коленчатого тела. Таким образом, в слуховой коре происходит как последовательная, так и параллельная обработка. Исследования с помощью электроэнцефалографического картирования и функционального магнитного резонанса представляют доказательства подобной же организации слуховой коры у человека, и эта область коры активирована даже во время чтения по губам!)
По аналогии с другими сенсорными участками коры можно ожидать, что кохлеотопическая карта А, будет подразделена на различные функциональные зоны. Если двигать электрод вдоль изочастотной полоски, большинство клеток либо возбуждаются звуком, воспринимаемым любым из ушей (ЕЕ), либо возбуждаются из одного уха и тормозятся из другого (EI). Эта бинауральная чувствительность закономерно меняется, и полагают, что ее значения образуют полосы, идущие под прямым углом к изочастотным контурам. Существуют также некоторые доказательства закономерных изменений кодирования интенсивности и ширины полосы в A1. Организация ЕЕ-EI может быть выведена из бинауральных взаимодействий, которые сначала возникают в ядрах олив, и приводит к предположению, что А, также картирована для слухового пространства, хотя в настоящее время это не доказано.
Обработка слуховых сигналов сложна. Важные с точки зрения поведения звуки должны быть извлечены из обширного и многообразного акустического окружения. Не только частотный состав, но также и временная последовательность входов должна быть проанализирована каким-то образом (например, проигрывание магнитофонной записи человеческой речи в обратную сторону воспринимается как тарабарщина). Кроме того, такая обработка у любого вида животных должна иметь дело не только с анализом окружающих звуков, но также со способами извлечения звука разными видами. У людей основные элементы речи, называемые фонемами, одинаковы для всех языков и являются звуками, которые первыми лопочут дети, прежде чем какие-то конкретные звуки будут выбраны, чтобы сочетать их в слова. Базовые звуки могут быть проанализированы как комбинации частотно-временных отношений, например, продолжительный компонент на 1 000 Гц сопровождается вторым, частотно-модулированным компонентом, начинающимся с 5 000 Гц и быстро снижающимся до 500 Гц. Компоненты называются формантами.
По аналогии со зрительной системой, которая содержит клетки, распознающие разрезы, углы, края и другие геометрические формы, мы могли бы ожидать, что обнаружим клетки высокого порядка в слуховой коре человека, которые реагируют на конкретные форманты, или, возможно, фонемы. Этот принцип, действительно, наблюдается у животных. Например, некоторые клетки в слуховой коре усатой летучей мыши реагируют на конкретные комбинации тонов постоянной и модулируемой частот, которые эквивалентны собственным звукам летучей мыши. В то же время, мы могли бы ожидать, что напрасно будет искать клетки, отвечающие за человеческие звуковые элементы в слуховой системе летучей мыши или других млекопитающих. Такие клетки высокого порядка, однако, все же обнаружены у птицы рода Mynah, которых обучают говорить. Кортикальные механизмы языкового кодирования могут быть обнаружены у таких видов, как мармозетки, которые используют ограниченный и стереотипный набор призывов ("слов") для общения.
Локализация звука
Чрезвычайная чувствительность и частотная избирательность слуховой системы была представлена у животных задолго до того, как развилась человеческая речь. В значительной степени тонкости слуховой системы возникли, чтобы улучшить способность организма определять местоположение звука в пространстве. Преимущества этого очевидны: сигналы, передающиеся на большие расстояния как звуковые волны, могут помочь обнаружить хищника или жертву при отсутствии зрительной и другой информации. Однако, в отличие от зрительной или соматосенсорной систем, звуковой нейроэпителий не может быть использован для кодирования места (локации), поскольку он приспособлен для тонотопического картирования. Вместо этого направление звука вычисляется на основе бинаурального сравнения временного хода и интенсивности сигналов, которое производится в центральной слуховой системе. Слуховой проводящий путь, поэтому, сложен и включает в себя многочисленные подкорковые синаптические переключения и многочисленные перекрещивания почти на каждом уровне.
Дорзальное кохлеарное ядро в основном предназначено для мензурального частотного анализа119) и обеспечивает относительно прямую, тонотопически организованную проекцию на контралатеральную область А1. Напротив, нейроны второго порядка в вентральном кохлеарном ядре проецируются как ипсилатерально, так и контралатерально на комплекс верхних олив в стволе мозга. Большинство нейронов в медиальной верхней оливе (MSO) возбуждаются стимуляцией любого из ушей (и, таким образом, соотносятся с ЕЕ-нейронами, но реагируют лучше всего, когда тон подается на оба уха с характерной задержкой, соответствующей достижению сначала одного уха, потом другого. Скорость звука в воздухе составляет 340 м/с, так что максимальная временная разница, создаваемая человеческой головой (около 18 см в диаметре), составляет 0,5 мс для звука, приходящего вдоль оси между обоими ушами, и значительно меньшие задержки для звуков, приходящих спереди. В дополнение к разнице во времени прибытия, постоянный источник звука вызывает фазовый сдвиг между двумя ушами.
Клетки латеральной верхней оливы (LSO) получают возбуждение ипсилатерального вентрального кохлеарного ядра. Клетки в контралатеральном вентральном кохлеарном ядре проецируются на противоположную сторону и образуют синапсы на медиальном ядре трапециевидного тела (MNTB). Клетки MNTB тормозят нейроны в LSO.
Таким образом, нейроны в LSO возбуждаются ипсилатеральным и подавляются контралатеральным звуком (El). Такое взаимодействие может быть полезным для выявления различий в интенсивности звука в двух ушах. На высоких частотах обнаруживается 10-кратная разница в частоте, для которой голова служит как эффективное затенение звука.
Различия как по фазе, так и по интенсивности изменяются как функция частоты. Для головы человека разница по фазе значительна на частотах ниже 2 кГц, тогда как отличия по интенсивности становятся более существенными на более высоких частотах. Психофизические исследования показали, что локализация осуществляется сопоставлением различий между двумя ушами по времени прихода сигнала и/или интенсивности входящего звука. Таким образом, если подаются щелчки через наушники с различными задержками, звук локализуется по направлению к уху, к которому он приходит раньше. Если щелчки подаются одновременно, но с разной интенсивностью, то звук локализуется с той стороны, где щелчок громче. Фактически сигналы локализации зависят в точности от частотного содержания. Голова и внешнее ухо отфильтровывают специфические компоненты частоты, в зависимости от места в пространстве, откуда исходит звук. Эти спектральные метки являются важными сигналами, которые используются для локализации звука. Люди могут различать интерауральную временную разницу всего в 5 мкс - очень высокое разрешение, если учесть, что длительность потенциала действия составляет примерно 1 мс; это показывает важность точной временной организации для слуховых функций.
Выводы
Соматосенсорные рецепторы в коже отвечают на разнообразные стимулы, такие как прикосновение, давление, вибрация и перемена температуры. Рецепторы различаются по скорости адаптации и по размеру рецептивного поля.
Адаптация сенсорных ответов зависит от механических свойств вспомогательных структур и от изменений в возбудимости. Быстро адаптирующиеся рецепторы "настроены" на более высокочастотную стимуляцию.
Центральные соматосенсорные проводящие пути соматотопически картированы. Соматотопическая карта коры подразделена на функциональные области, отражающие специфичность стимула периферических рецепторов.
Рецептивные поля нейронов в соматосенсорной коре организованы по принципу "центр - окружение".
Механическая настройка базилярной мембраны у млекопитающих обеспечивает частотную избирательность ответов слуховых афферентов. Потенциалзависимая подвижность внешних волосковых клеток вносит вклад в акустическую чувствительность улитки млекопитающих.
Электрическая настройка обеспечивает избирательность волосковых клеток низших позвоночных. Взаимодействие потенциалзависимых кальциевых каналов с кальций-активируемыми калиевыми каналами приводит к усилению ответа мембранного потенциала на частоте, равной характеристической частоте звука для каждой волосковой клетки.
Волосковые клетки улитки подвергаются эфферентному торможению холинергическими нейронами ствола мозга. Эфферентное торможение изменяет чувствительность и настройку афферентов улитки.
Центральные слуховые проводящие пути, включая кору, тонотопически картированы. Свойства ответов клеток в слуховой коре сложны и включают бинауральные взаимодействия и зависимость от временных комбинаций тонов.
Бинауральные комбинации интенсивности и временных параметров звука используются для того, чтобы вычислять местоположение источника звука в пространстве. Эти вычисления выполняются синаптическими связями в ядрах верхней оливы.
Литература:
1. Fettiplace, R., and Fuchs, P. A. 1999. Mechanisms of hair cell tuning. Anna. Kev. Physiol.61: 809-834.
2. Kaas, J. H., Hackeit, Т.Д., and Tramo, M. J. 1999. Auditory processing in primate cerebral cortex. Curr. Opin. Neurobiol.9: 164-170.
3. Mountcastle, V. B. 1995. The parietal system and some higher brain functions. Cerebral Cortex 5: 377-390.
4. Mountcastle, V. B. 1997. The columnar organization of the neocortex. Brain 120: 701-722.
5. Patuzzi, R. 19%. Cochlear micromechanics and macromechanics. In P. Dallos, A. N. Popper, and R. R. Fay (eds), The Cochlea. Springer, New York, pp.186-257.
© 2010 Интернет База Рефератов