
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Реферат: Эколого-фитоценотическая структура бриокомпонента лесной растительности Республики Башкортостан
Реферат: Эколого-фитоценотическая структура бриокомпонента лесной растительности Республики Башкортостан
На правах рукописи
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
доктора биологических наук
Эколого-фитоценотическая структура бриокомпонента лесной растительности Республики Башкортостан
Специальность 03.02.01 – ботаника
БАИШЕВА ЭЛЬВИРА ЗАКИРЬЯНОВНА
УФА – 2010
Работа выполнена в Лаборатории геоботаники и охраны растительности Учреждения РАН Института биологии Уфимского научного центра РАН
| Научный консультант |
Миркин Борис Михайлович доктор биологических наук, профессор |
| Официальные оппоненты: |
Игнатов Михаил Станиславович доктор биологических наук |
|
Ермаков Николай Борисович доктор биологических наук |
|
|
Ишбирдин Айрат Римович доктор биологических наук, профессор |
|
| Ведущая организация |
Учреждение Российской академии наук Полярно-альпийский ботанический сад-институт им. Н.А. Аврорина Кольского научного центра РАН |
Защита состоится «11» ноября 2010 г. в 1400 часов на заседании диссертационного совета Д 212.013.11 при ГОУ ВПО «Башкирский государственный университет» по адресу: 450074, г. Уфа, ул. Заки Валиди, 32, биологический факультет, ауд. 332; факс (347) 273-67-78, e-mail: disbiobsu@mail.ru.
Официальный сайт БашГУ: http://www.bashedu.ru
С диссертацией можно ознакомиться в библиотеке ГОУ ВПО «Башкирский государственный университет».
Ученый секретарь диссертационного совета, д.б.н. М.Ю. Шарипова
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы исследования. Мохообразные (Bryophyta, Marchantiophyta и Anthocerotophyta) – обособленная и древняя группа высших растений, которые распространены на всех континентах и характеризуются высоким морфологическим и таксономическим разнообразием [Абрамов, Абрамова, 1978; Потемкин, 2007]. По видовому богатству бриофиты стоят на втором месте после цветковых растений [Wilson, Peter, 1988], но изучены гораздо хуже. Роль мохообразных в формировании флоры и растительности часто недооценивается, а их таксономическое разнообразие в пределах ландшафтов и разных типов растительности до настоящего времени остается одним из наименее исследованных вопросов экологии растений [Newmaster et al., 2003]. Мохообразные часто используются в качестве индикаторов состояния среды обитания [Vitt, Belland, 1997; Hokkanen, 2004 и др.], в том числе при контроле загрязнения воды и атмосферы [Burton, 1990; Tyler, 1990; Zechmeister, 1998 и др.].
На территории Республики Башкортостан (РБ) мохообразные входят в состав всех типов растительности и вносят вклад в формирование биологического разнообразия региона: каждый пятый вид высших растений на территории республики – мох или печеночник. Тем не менее, бриофиты исследованы в меньшей степени, чем сосудистые растения, что снижает эффективность охраны их разнообразия и затрудняет использование этих растений при мониторинге состояния экосистем. Особую актуальность исследования мохообразных приобретают в современных условиях высокой степени антропогенной трансформации растительного покрова региона.
Цель исследования – выявление и анализ флоры мохообразных лесной, лесостепной зон РБ и их горных аналогов, изучение закономерностей распределения бриофитов в разных типах лесной растительности РБ, классификация бриосообществ, выявление видов-индикаторов лесных сообществ с высокой природоохранной ценностью, а также обоснование подходов к охране мохообразных.
Задачи исследования:
1. Характеристика и сравнительный анализ бриоценофлор ассоциаций и союзов коренных зональных лесов РБ и их горных аналогов.
2. Анализ экологической обусловленности половых типов и жизненных стратегий мохообразных.
3. Выявление видов-индикаторов сообществ биологически ценных лесов РБ.
4. Разработка бриосинтаксономии (эколого-флористической классификации) эпифитных, эпиксильных и прибрежно-водных бриосообществ РБ.
5. Инвентаризация видового состава флоры мохообразных лесной и лесостепной зон РБ, а также их горных аналогов.
6. Анализ состава редких видов мохообразных РБ и разработка рекомендаций по их охране.
Основные положения диссертации, выносимые на защиту:
1. Невысокий уровень флористического сходства лесных бриоценофлор РБ, отражающий преобладание видов с низким постоянством в сообществах.
2. Использование различий спектров половых типов и жизненных стратегий мохообразных разных субстратных групп для экологического мониторинга состояния лесных экосистем и выявления участков биологически ценных лесов.
3. Высокое сходство прибрежно-водных бриосообществ РБ с центрально-европейскими аналогами и существенные отличия эпифитных и эпиксильных сообществ, обедненный флористический состав которых отражает континентальность климата Южного Урала.
4. Необходимость специальных мер по охране мохообразных РБ в зонах интенсивной рекреации и лесах хозяйственного использования, в связи с тем, что охрана в системе ООПТ не является достаточным условием для сохранения их биоразнобразия.
Научная новизна. В лесной и лесостепной зонах РБ выявлено 365 видов листостебельных мхов и 91 вид печеночников. Впервые в регионе найдены 41 вид листостебельных мхов и 38 печеночников. Анализ бриоценофлор 42 ассоциаций коренных зональных лесов РБ и их горных аналогов показал, что для большинства мохообразных характерно низкое постоянство в сообществах. Виды эпифитно-эпиксильного комплекса имеют преимущественно сквозное распространение в лесах разных типов, менее эвритопные эпигейные бриофиты могут быть использованы в качестве диагностических видов при выделении низших единиц классификации лесной растительности. Мохообразные разной субстратной приуроченности различаются по спектрам половых типов и жизненных стратегий, что должно учитываться при мониторинге лесных экосистем. Впервые выявлены группы мохообразных-индикаторов биологически ценных лесов региона. Впервые разработана эколого-флористическая классификация прибрежно-водных, эпифитных и эпиксильных бриосообществ РБ, описано 5 новых ассоциаций, выявлены различия южно-уральских сообществ с аналогами из Западной и Центральной Европы.
Научно-практическая значимость работы. Результаты работы были использованы при составлении “Cписка мхов Восточной Европы и Северной Азии” [Ignatov et al., 2006] и Красной книги РБ [2002, 2007]. При выполнении исследований была проведена инвентаризация бриофлор основных ООПТ РБ: Южно-Уральского государственного природного заповедника, заповедника «Шульган-Таш», национального парка «Башкирия», проектируемого природного парка «Инзер» и др. Фонд Гербария Института биологии УНЦ РАН (UFA) пополнился 4500 образцов мохообразных. Полученные результаты использованы при разработке лекций, прочитанных автором студентам-биологам Башкирского государственного педагогического университета им. Акмуллы.
Связь работы с плановыми исследованиями и научными программами. Исследования флоры мохообразных РБ проводились при выполнении планов НИР Лаборатории геоботаники и охраны растительности Института биологии УНЦ РАН: «Синтаксономия как основа мониторинга и охраны биологического разнообразия растительного мира Южного Урала» (1998-2002), «Разработка теоретических основ системы охраны биологического разнообразия растений Южного Урала на уровне видов и сообществ» (2003-2005), «Анализ состояния системы охраны флоры и растительности Южно-Уральского региона и обоснование прогноза ее дальнейшего развития» (2006-2008), «Синтаксономический анализ растительности болот Южно-Уральского региона, оценка состояния популяций редких видов с разработкой теоретических и практических основ их охраны» (2009-2012), при поддержке грантов РФФИ № 97-04-48017, № 04-04-49269-а, № 07-04-00030-а; 10-04-00534-а, грантов РФФИ-Агидель № 02-04-97914, № 02-04-97927; программ ГНТП АН РБ № 17-8-1/1, № 19-102-05, Программы Президиума РАН «Биологическое разнообразие» (подпрограмма «Разнообразие и мониторинг лесных экосистем России»).
Декларация личного участия. Автором лично определены цели и задачи исследований, в течение 20 лет проведено более 40 экспедиционных выездов в различные районы РБ, в ходе которых была собрана обширная коллекция образцов мохообразных. Выполнено более 600 полных геоботанических описаний бриосообществ РБ, разработана синтаксономия бриосообществ республики, сформирована и пополняется коллекция мохообразных в Гербарии Института биологии УНЦ РАН (UFA). Определены образцы мохообразных к 1500 полным геоботаническим описаниям лесов РБ, проведены инвентаризация и эколого-ценотический анализ бриоценофлор лесов РБ, а также их сравнение с данными по бриофлоре лесных сообществ из других регионов России и стран Европы. Выявлены редкие и нуждающиеся в охране виды, разработаны рекомендации по их охране, ведется соответствующий раздел в «Красной книге РБ» [2007].
Апробация работы. Материалы диссертации были представлены на 33 конференциях, наиболее важные из которых: науч.-практ. конф. «Леса Башкортостана: современное состояние и перспективы» (Уфа, 1997), Межд. науч. конф. «Микология и криптогамная ботаника в России: традиции и современность» (Санкт-Петербург, 2000), Межд. науч. конф. «Биоразнообразие и биоресурсы Урала и сопредельных территорий» (Оренбург, 2001, 2006, 2008), науч. конф. «Проблемы изучения адвентивной и синантропной флоры в регионах СНГ» (Тула, 2003), Всеросс. науч.-практ. конф. «Уралэкология. Природные ресурсы – 2005» (Уфа, 2005), Всеросс. науч.-практ. конф. «Природная и антропогенная динамика наземных экосистем» (Иркутск, 2005), Межд. науч. конф. «Вопросы общей ботаники: традиции и перспективы» (Казань, 2006), II Всеросс. конф. «Научные аспекты экологических проблем России» (Москва, 2006), III Межд. науч.-техн. конф. «Наука, образование, производство в решении экологических проблем» (Уфа, 2006), III Всеросс. школа-конф. «Актуальные проблемы геоботаники» (Петрозаводск, 2007), 17th Intern. Workshop «European Vegetation Survey. Using phytosociological data to address ecological questions» (Brno, 2008), II Межд. научно-практ. конф. «Природное наследие России в 21 веке» (Уфа, 2008), Всеросс. конф. «Фундаментальные и прикладные проблемы ботаники в начале XXI века» (Петрозаводск, 2008), III Всеросс. науч. конф. «Принципы и способы сохранения биоразнообразия» (Пущино, 2008), Росс. науч. конф. «Раритеты флоры Волжского бассейна» (Тольятти, 2009), Межд. науч. конф. «Растительность Восточной Европы: классификация, экология и охрана» (Брянск, 2009), Межд. науч.-практ. конф. «Актуальные проблемы дендроэкологии и адаптации растений» (Уфа, 2009).
Публикации. По теме диссертации опубликовано 64 работы, в том числе главы в 3 монографиях, 10 статей в научных журналах, рекомендованных ВАК МОН РФ для защиты докторских диссертаций, 10 статей в центральных российских изданиях, 9 статей в крупных региональных изданиях.
Структура и объем работы. Диссертация состоит из введения, 8 глав, выводов, списка литературы и приложения. Объем работы 320 страниц машинописного текста, в том числе 8 таблиц, 11 рисунков. Приложение включает список видов, 10 таблиц, 1 рисунок и составляет 107 страниц. В списке литературы 456 наименований, в том числе 213 иностранных авторов.
Глава 1. Разнообразие мохообразных естественных экосистем: подходы к изучению и особенности охраны
Рассмотрены подходы, используемые при изучении разнообразия и эколого-ценотической структуры бриокомпонента экосистем, в том числе: традиционные методы флористического обследования [Шляков, 1961; Ignatova et al., 1995; Кривошапкин, 1998; Бойко, 1999; Афонина, 2000; Потемкин, 2002; Чернядьева, Потемкин, 2003; Tsegmed, 2003; Хмелев, Попова, 1988; Попова, 1998 и др.]; методы учета на пробных площадях [Slack 1977; Söderström, 1988; Gustafsson, Hallingbäck, 1988; Økland et al., 1990; Lesica et al. 1991; Frego, Carleton, 1995; Newmaster, Bell, 2002; Newmaster et al., 2005 и др.]; ординационные методы анализа растительности [Bradfield, Scagel, 1984; Laine, Vahna-Majamaa, 1992; Ganeva, 1996; Hokkanen, 2004]; классификация синузий [Gams, 1930; Lippmaa, 1939; Улычна, 1989; Арискина, 1962; Лавренко, 1962; Трасс, 1964; Du Rietz, 1965; Сукачев, 1975; Корчагин, 1976; Гудошников, 1978; Бойко, 1978; Норин, 1979; Отнюкова, 1985, 1986 и др.] и эколого-флористическая классификация бриосообществ [Gams, 1927; Barkman, 1958; Hübschmann, 1986; Marstaller, 2006].
Отмечается, что на ландшафтном уровне распространение мохообразных, в основном, определяется наличием и качеством подходящих для них субстратов и местообитаний [van Zanten, Pocs, 1981; Söderström, 1989; Вьюнова, 1989; Miles, Longton, 1992; Vitt, Belland, 1997; Pharo, Beattie, 2002; Pharo et al., 2004; Дьяченко, 1999 и др.], что обуславливает некоторую автономность представителей этой группы от окружающей растительности [Бойко, 1978; Улычна и др., 1989; Коротков, Морозова, 1989; Newmaster, Bell, 2002; Newmaster et al., 2005; Økland et al., 1990].
Для эффективного сохранения биоразнообразия бриофитов необходим мониторинг за его состоянием. Важную роль при сравнении и обобщении результатов мониторинговых исследований может играть использование эколого-флористической классификации растительности [Миркин и др., 2004]. Мохообразные, как правило, формируют пространственно обособленный компонент фитоценоза, со своей структурой, составом жизненных форм и взаимосвязями между видами. В соответствии с подходом Браун-Бланке, группировки мохообразных считаются малыми сообществами и классифицируются как самостоятельные ассоциации, объединенные в союзы, порядки и классы [Hübschmann, 1986; Marstaller, 1993, 2006]. В России и странах СНГ бриосинтаксономия находится на начальном этапе. Существуют отдельные работы, посвященные классификации бриосообществ Южного Урала [Baisheva et al., 1994; Baisheva, 1995, 2000; Баишева и др., 2004]; Сибири [Писаренко, 1997; Pisarenko, 1999]; Нижегородской области [Шестакова, 2005]; Юго-Западного Нечерноземья России [Анищенко, 2001, 2007], Левобережной лесостепи Украины [Гапон, Баïшева. 2002]. В настоящее время на территории России описано 54 ассоциации, принадлежащие к 14 союзам, 10 порядкам и 6 классам растительности мохообразных [Баишева, 2007].
Рассмотрены типы эколого-фитоценотических стратегий мохообразных и возможности их применения при оценке экологического режима местообитаний и степени антропогенного воздействия на природные экосистемы [Slack, 1977; During, 1979, 1992; Furness, Grime, 1982; Grime et al., 1990; Ахминова, 1983; Rincon, Grime, 1988; Thiers, 1988; Рыковский, 1989; Gradstein, Pocs, 1989; Бойко, 1990; Longton, Hedderson, 2000; Андреева, 2002; Kürschner, 2000, 2004]. Обсуждается влияние на разнообразие мохообразных различных нарушений (пожаров и рубок леса) [Southorn, 1976; During, 1979; During, Tooren, 1990; Gustafsson, Hallingbäck 1988; Jonsson, Essen 1990; Clément, Touffet, 1990; Andersson, Hytteborn 1991; Heras-Ibáñez et al. 1990, 1991, 1992; Rozé, 1993; Söderström et al. 1992; Söderström 1995; Esposito et al., 1999; Newmaster et al. 1999, Bell, Newmaster 2002; Newmaster et al., 2003; Sim-Sim et al., 2004 и др.], а также результаты изучения банка диаспор мохообразных и его роли в восстановительных сукцессиях растительности [During, ter Horst, 1983; During et al., 1987; Smith, 1987; Jonsson, Esseen, 1990; Jonsson, 1993; Frego, 1996].
Особое внимание уделено особенностям бриокомпонента лесных экосистем [Курский, 1909; Grebe, 1917; Сукачев, 1927; Буш, Аболинь, 1968; Улична, 1970; Ахминова, 1970, 1983; Furness, Grime, 1982; Rincón, Grime, 1989; Rambo, Muir, 1998; Бардунов, 1965, 1974, 1992; Бардунов, Васильев, 2005; Черепанова, Ипатов, 2003; Økland, 1994; Vellak et al., 2003; Шестакова, 2004 и др.], особенно, старовозрастных лесов [La Roi, Stringer, 1976; Johnson 1981; Foster 1985; Edwards 1986; Söderström 1988; Gustafsson, Hallingbäck 1988; Hallingbäck, Weibull, 1996; Carleton 1990; Frego, Carleton 1995; Laaka 1992; Crites, Dale 1995; Esseen et al., 1992; Trass et al., 1999; Vellak, Paal, 1999; Maksimov et al., 2003; Newmaster et al., 2003]. Целенаправленные исследования бриокомпонента лесных сообществ (инвентаризация и сравнительный анализ бриоценофлор) проводились в немногих регионах России [Гудошников, 1986; Хмелев, Попова, 1988; Игнатов, Игнатова, 1990; Мордвинов, Благовещенский, 1995; Дегтева и др., 2001; Шубина, Железнова, 2002; Писаренко, Таран, 2001; Чуракова, 2002; Шестакова, 2004, 2005; Слука, 2004; Дегтярева, 2004; Нешатаева и др., 2004; Анищенко, 2007; Дулин, 2007 и нек. др.]. За исключением экосистем бореальных лесов, состав мохообразных в лесах большинства регионов России остается малоизученным. Мохообразные хорошо приспособлены к обитанию в лесной среде и освоили широкий спектр экотопов. Состав бриофитов способен отражать изменения экологического режима сообществ, степень атмосферного загрязнения и уровень антропогенной дигрессии лесных экосистем. Тем не менее, в отечественном лесном хозяйстве и природоохранной практике прикладные аспекты использования мохообразных пока не получили должного научного обоснования и развития [Малышева, 1998; Дегтярева, 2004].
Обсуждаются вопросы организации охраны мохообразных, особенности экологического менеджмента и выявления видов, нуждающихся в охране [Игнатов, Игнатова, 1990; Белкина и др., 1991; Red Data Book…, 1995; Söderström, 1995; Hedenäs, 1996; Hodgetts, 1996; Neitlich, McCune 1997; Красная книга Республики Коми, 1998; Ingerpuu, Vellak, 2000; Максимов, 2000; Longton, Hedderson, 2000; Bisang, Hedenäs, 2000; Urmi, Schnyder, 2000; Чернядьева, 2001; Vanderpoorten et al., 2001; Boudreault et al., 2002; Heinlen, Vitt, 2003; Manzke, Wentzel, 2003; Jukoniené, 2005].
Глава 2. МАТЕРИАЛЫ И МЕТОДЫ
Основой для написания работы послужили результаты обработки коллекций листостебельных мхов и печеночников, собранных во время полевых исследований 1991-2009 гг. Флористическими исследованиями охвачено более 400 пунктов сбора во всех природных зонах республики, собрано около 20000 образцов мохообразных. Кроме собственных коллекций были обработаны сборы Г.В. Попова, А.А. Мулдашева, В.Б. Мартыненко, П.С. Широких, А.И. Соломеща, И.Н. Григорьева, А.Р. Ишбирдина и др. Обработанные коллекции хранятся в Гербарии Института биологии УНЦ РАН (UFA), отдельные образцы переданы в гербарий Московского государственного университета (MW) и бриологический гербарий БИН РАН (LE). При определении материала использовался общепринятый анатомо-морфологический метод. В сложных случаях проводились консультации с Е.А. Игнатовой (МГУ), М.С. Игнатовым (ГБС РАН), А.Д. Потемкиным (БИН РАН), В.И. Золотовым (ГБС РАН), А.М. Максимовым (ИБ КНЦ РАН). При анализе бриофлоры использовались ботанико-географический и сравнительно-флористический методы.
Материал для изучения лесных бриоценофлор собран автором и сотрудниками лаборатории геоботаники и охраны растительности Института биологии УНЦ РАН при геоботаническом обследовании растительности РБ в 1989-2008 гг. Эти данные характеризуют бриокомпонент 42 ассоциаций лесной растительности, выделенных на основе обработки 1433 полных геоботанических описаний на площадках размером 400-1000 м². В пределах пробных площадей проводилось выявление видового состава мохообразных на всех типах субстрата: стволах и комлях деревьев, гнилой древесине, валеже, почве, скальных выходах и пр. Для напочвенных видов мхов отмечалось обилие по шкале Браун-Бланке [Braun-Blanquet, 1964]. Для анализа взаимосвязи бриокомпонента с параметрами растительных сообществ и условиями местообитаний использованы методы градиентного анализа: канонический анализ соответствий (СCA) и анализ соответствий с удаленным трендом (DCA).
Объединения бриофлористических списков однотипных экологически и структурно сходных фитоценозов могут рассматриваться как парциальные ценофлоры [Юрцев, 1994] или бриокомпоненты ценофлор [Седельников, 1987]. Соответственно, к ним применимы методы флористического анализа. В работе использованы коэффициенты Жаккара, Съеренсена-Чекановского и меры включения.
Классификация бриосообществ выполнена в соответствии с подходом Браун-Бланке [Braun-Blanquet, 1964; Westhoff, van den Maarel, 1978; Миркин и др., 1989; Dierschke, 1994] на основе обработки 1200 описаний. Для описаний эпифитных и эпиксильных бриосообществ выбирались наиболее типичные (часто повторяющиеся) участки мохового покрова на стволах деревьев и гнилой древесине. При этом особое внимание уделялось экологической гомогенности микроместообитаний (увлажнению, освещению, экспозиции и пр. условиям) в пределах пробной площадки. Размер площадки варьировал от 1 до 8 дм². При изучении водных и прибрежно-водных бриосообществ выбирались участки с однородным растительным покровом в русле и по берегам ручьев и малых рек. Размер площадок варьировал от 2 дм² до 6 м², в зависимости от величины участков с однородными условиями (камни в русле, отмели, песчано-глинистые наносы, подтопленные участки по берегам и др.). В полевых условиях оценивался предварительный состав видов, окончательный список устанавливался при камеральной обработке после определения видов с привлечением микроскопии. При разработке эколого-флористической классификации сообществ мохообразных использованы классификационные системы из Центральной и Западной Европы [Barkman, 1958; Hübschmann, 1986; Marstaller, 1993, 2006 и др.].
Обработка данных осуществлена с помощью программ TURBOVEG, MEGATAB [Hennekens, Schaminee, 2001], CANOCO 4.5 [ter Braak, Šmilauer 2002], программного модуля “GRAPHS” [Новаковский, 2004].
Названия видов листостебельных мхов даны в соответствии с «Check-list of mosses of East Europe and North Asia» [Ignatov et al., 2006], названия печеночников – по «Аннотированному списку печеночников и антоцеротовых России» [Потемкин, Софронова, 2009] и «Списку печеночников (Marchantiophyta) России» [Константинова и др., 2009]. Названия синтаксонов соответствуют требованиям «Международного кодекса фитосоциологической номенклатуры» [Weber et al., 2000; Вебер и др., 2005].
ГЛАВА 3. ХАРАКТЕРИСТИКА ПРИРОДНЫХ УСЛОВИЙ РЕСПУБЛИКИ БАШКОРТОСТАН
В главе охарактеризованы геология и рельеф, климат, гидрография и гидрология, почвы и почвообразующие породы, дана общая характеристика растительности и природных районов РБ [Крашенинников, Кучеровская-Рожанец, 1941; Физико-географическое…, 1964; Жудова, 1966; Тайчинов, 1973; Агроклиматические ресурсы..., 1976; Попов, 1980; Почвы…, 1995; Мукатанов, 1992; 2002; Атлас…, 2005; Реестр, 2006 и др.].
Башкортостан расположен между 51°34' 56°31' с.ш. и 53°08' – 60°00' в.д., площадь 143,6 тыс. км2. Территория республики находится в зоне сочленения Восточно-Европейской (Русской) платформы и Уральской складчатой области, подразделяется на три крупные геоморфологические области: равнины Южного (Башкирского) Предуралья, горы Южного Урала и равнины Южного (Башкирского) Зауралья. На западном макросклоне Южного Урала преобладают карбонатные почвообразующие породы, а в центрально-возвышенной части и на восточном макросклоне – магматические и метаморфические породы.
Климат континентальный, с умеренно теплым, иногда жарким летом и продолжительной умеренно холодной зимой. Уральские горы являются преградой на пути влажных воздушных масс Атлантики, в результате чего климат на западном макросклоне и в его предгорьях более влажный и теплый, а на восточном – более континентальный.
Таблица 1 Основные климатические показатели РБ
| Климатические показатели |
Башкирское Предуралье |
Южный Урал | Башкирское Зауралье |
| Среднегодовая t воздуха, °C | +2,5 - +3,5 | +0,5 - +2,0 | +1,5 - +2,0 |
| Средняя t воздуха. Январь, °C | –14,5 - –15,5 | –15,5 - –17 | –15,5 - –16 |
| Средняя t воздуха. Июль, °C | +18,5 - +19,5 | +16,5 - +17,5 | +18,0 - +19,5 |
|
Продолжительность безморозного периода, дни |
55 – 100 | 50 – 80 | 65 – 120 |
| Среднегодовое количество осадков, мм | 450 – 500 | 600 – 700 | 350 – 400 |
Растительный покров РБ характеризуется высоким разнообразием. Леса занимают 39,2 % территории республики (6,2 млн га). На территории РБ представлены зоны широколиственных и бореальных лесов, а также их горные аналоги (на долю горных лесов приходится 4,5 млн га). В системе эколого-флористической классификации обследованные типы леса отнесены к 3 классам, 4 порядкам, 9 союзам, 10 подсоюзам и 48 ассоциациям [Мартыненко, 2009].
Естественная травянистая растительность занимает около 16 % территории РБ. Основная часть этой площади занята горными каменистыми степями, также представлены равнинные степи и луга, низинные травяные болота, фрагменты горных тундр, субальпийских и альпийских лугов и верховых болот. Луга, как правило, имеют вторичный характер и формируются на месте вырубленных лесов при регулярном сенокошении или выпасе. Болота занимают менее 0,4 % территории РБ, в основном, сосредоточены в северо-западных и горно-лесных районах. В высокогорьях (г. Иремель, Яман-Тау, Большой Шелом и др.) встречаются сообщества горных тундр. Значительную площадь занимают сегетальные и рудеральные сообщества [Научно-обоснованные…, 1990].
ГЛАВА 4. История изучения флоры мохообразных РБ
Сведения о первых находках мхов на современной территории РБ немногочисленны. Первым представителем мохообразных, который указан для Уфимской и Оренбургской губерний был Polytrichum commune [Georgi, 1775], а к 1880 г. для Южного Урала было известно всего 5 видов мхов [Bunge, 1854; Meinshausen, 1859]. В конце XIX – начале XX веков значительный вклад в изучение мохообразных Южного Урала внесли Ю.К. Шелль [1880, 1883], О.А. и Б.А. Федченко [Федченко, Федченко, 1894], Э.В. Цикендрат [Zickendrath, 1894, 1900], В.И.Талиев [1903]. Большой список мохообразных территории Уфы и ее окрестностей, содержащий 155 видов и многочисленные внутривидовые таксоны, опубликовал чешский бриолог Й. Подпера [Podpěra, 1921]. Некоторые сведения о сфагновых мхах Южного Урала приведены Д.А. Герасимовым [1926, 1931], о печеночниках – З.Н. Смирновой [Смирнова, 1928, 1931].
В годы Великой Отечественной войны на территории Башкирской АССР находились в эвакуации и работали ученые из Института ботаники АН Украины. По результатам этих исследований А.Ф. Бачурина [1946] опубликовала список 65 видов листостебельных мхов Башкирского Предуралья, Д.К. Зеров [1947] обобщил сведения о 27 видах сфагновых мхов Южного Урала и Башкирского Предуралья.
В 1945, 1946 и 1948 гг. сотрудником БИН АН СССР Е.А. Селивановой-Городковой была обследована территория Башкирского государственного заповедника, для которого был составлен список из 145 видов листостебельных мхов и 24 печеночников [Селиванова-Городкова, 1956; Селиванова-Городкова, Шляков, 1956]. Отмечалось, что находки неморальных бриофитов подтверждают ценность Южного Урала как рефугиума, где сохранились представители третичной флоры. В качестве особенностей бриофлоры Южного Урала рассматривались отсутствие эндемиков, а также наличие видов, находящихся на границе своего ареала. В общей сложности в этих работах указано 57 видов мохообразных, новых для Южного Урала [Селиванова-Городкова, 1956 а]. Некоторые наиболее массовые виды мохообразных указаны в геоботанических работах [Герасимов, 1926, 1931; Носков, 1931, Бобров, 1928; Тюлина, 1931; Генкель, Осташева, 1933; Котов, 1947, 1959; Крашенинников, Кучеровская-Рожанец, 1941; Крашенинников, Васильев, 1949; Брадис, 1951, 1959; Горчаковский, 1954, 1972, 1975, Шарафутдинов, 1983 и др.]. Кроме того, были опубликованы работы, посвященные изучению ископаемых видов мхов Башкирии [Абрамов, 1965] и хромосомных чисел листостебельных мхов Южного Урала [Данилкив и др., 1984]. Первое обобщение сведений по листостебельным мхам Башкирии было выполнено Е.А. и М.С. Игнатовыми, которые опубликовали список из 302 видов [Игнатова, Игнатов, 1993]. Позже перечень листостебельных мхов РБ пополнился еще 65 видами в результате новых находок [Баишева, 2002; Баишева, Игнатова, 2009] и таксономических ревизий [Ignatov et al., 2006]. Первая сводка по гепатикофлоре республики насчитывала 67 видов [Баишева, Потемкин, 1998], позже она была дополнена еще 24 видами [Шляков, 1998; Potemkin A.L., Kalinauskaite, 2008; Потемкин, Баишева, 2009; Баишева, Жигунова, 2007; Баишева, Широких, 2008]. На настоящий момент бриофлора РБ насчитывает 367 видов листостебельных мхов и 91 вид печеночников.
ГЛАВА 5. Обзор бриоценофлор лесных сообществ РБ
В главе приведена подробная характеристика бриокомпонента 42 ассоциаций лесов РБ. Представлены характеризующие таблицы, отражающие постоянство видов в ассоциациях, с помощью методов сравнительной флористики проведено сравнение лесных бриоценофлор РБ с данными по лесам других регионов: Республики Коми [Дегтева и др., 2001], Салаирского Кряжа [Писаренко, 1997], Алтая и Саян [Гудошников, 1986], бассейна Среднего Дона [Хмелев, Попова, 1988], лесостепных районов Украины [Гапон, 1992; Бойко 1975, 1999], Ленинградской [Курбатова, 2002], Ульяновской [Мордвинов, Благовещенский, 1995], Волгоградской [Сурагина, 2001] Тульской [Абрамова, Курнаев, 1977] областей и др.
При анализе бриокомпонента синтаксонов была использована классификация лесной растительности РБ, разработанная В.Б. Мартыненко [2009]. Приведем краткую характеристику исследованных союзов:
Класс Querco-Fagetea Br.-Bl. et Vlieger in Vlieger 1937 (неморальные широколиственные и хвойно-широколиственные леса европейского типа):
1. союз Alnion incanae Pawłowski, Sokołowski et Wallisch 1928 (пойменные леса с ольхой серой, черемухой, вязом, формирующиеся в долинах рек и ручьев на плодородных, затапливаемых во время весенних паводков, хорошо увлажненных, но не заболоченных почвах);
2. союз Lathyro-Quercion roboris Solomeshch et al. 1989 (ксеромезофитные злаково-разнотравные дубовые леса, развивающиеся, как правило, на границе леса со степью или с остепненными лугами);
3. союз Aconito septentrionalis-Tilion cordatae Solomeshch et al. 1993 (широкотравные леса с липой, кленом, дубом, распространенные на западном макросклоне Южного Урала и в южной оконечности его центральной части. Они приурочены к серым лесным почвам и занимают плакорные местообитания, надпойменные террасы, а также склоны гор различной крутизны и экспозиции. В качестве подсоюза в данный союз включены мезофитные и ксеромезофитные светлохвойно-широколиственные леса с разреженным травяным ярусом из неморального широкотравья, вейниково-разнотравного и бореально-мелкотравного комплексов);
4. союз Aconito septentrionalis-Piceion obovatae Solomeshch et al. ex Martynenko et al. 2008 (неморальнотравные темнохвойные и темнохвойно-широколиственные леса, встречающиеся на серых и светло-серых лесных почвах в зонах южной и горной тайги).
Класс Brachypodio pinnati-Betuletea pendulae Ermakov, Koroljuk et Latchinsky 1991 (светлохвойные и мелколиственно-светлохвойные гемибореальные леса сибирского типа):
5. союз Caragano fruticis-Pinion sylvestris Solomeshch et al. 2002 (остепненные сосновые и сосново-лиственничные леса Южного Урала с подлеском из степных кустарников и травяным ярусом из луговостепных, степных и петрофитных видов. Они растут на щебнистых перегнойно-карбонатных горно-лесных почвах на крутых склонах, сложенных известняками и доломитами);
6. союз Veronico teucrii-Pinion sylvestris Ermakov et al. 2000 (мезоксерофитные березово-сосновые и березовые травяные леса лесостепной зоны и средних частей склонов гор Южного Урала);
7. союз Trollio europaea-Pinion sylvestris Fedorov ex Ermakov et al. 2000 (мезофитные и гигромезофитные вейниково-широкотравные леса с сосной, лиственницей, березой, развивающиеся на относительно плодородных и хорошо обеспеченных влагой почвах в средних и нижних частях склонов).
Класс Vaccinio-Piceetea Br.-Bl. in Br.-Bl., Sissingh et Vlieger 1939 (таежные бореальные леса):
8. союз Dicrano-Pinion (Libbert 1933) Matuszkiewicz 1962 (ксеромезофитные зеленомошно-лишайниковые сосняки и лиственничники, приуроченные к кислым слаборазвитым почвам нижних, средних, реже - верхних частей склонов);
9. союз Piceion excelsae Pawłowski, Sokołowski et Wallisch 1928 (мезофитные и гигромезофитные темнохвойные зеленомошные и крупнотравно-зеленомошные леса).
Таксономический состав
В изученных лесных сообществах выявлено 212 видов мохообразных (180 видов листостебельных мхов и 32 - печеночников). Печеночники представлены 22 родами и 16 семействами, мхи – 95 родами и 37 семействами. Некоторые показатели бриоценофлор союзов приведены в таблице 2.
Ведущие семейства: Brachytheciaceae (21 вид), Dicranaceae (15), Amblystegiaceae (14), Mniaceae (12), Scapaniaceae s.l. (12), Grimmiaceae (10), Polytrichaceae (10), Pottiaceae (9), Bryaceae (8), Plagiotheciaceae (8), Pylaisiaceae (7), Sphagnaceae (6), Hylocomiaceae (6). Ведущие роды: Dicranum (13 видов), Brachythecium (9), Bryum (7), Sphagnum (6), Grimmia (6), Plagiomnium (5). Среднее число видов в роде 1,4, среднее число видов в семействе – 2,6.
Таблица 2 Общая характеристика бриоценофлор лесных союзов РБ
| Класс растительности | Querco-Fagetea | Brachypodio-Betuletea | Vaccinio-Piceetea | ||||||
| Число бриофитов | 160 | 106 | 151 | ||||||
| Номер союза | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| Число описаний | 96 | 123 | 346 | 195 | 88 | 101 | 183 | 120 | 184 |
| Число бриофитов | 71 | 63 | 104 | 121 | 80 | 63 | 71 | 74 | 138 |
| Число родов | 54 | 47 | 70 | 86 | 57 | 43 | 54 | 52 | 87 |
| Число семейств | 31 | 28 | 35 | 45 | 31 | 25 | 32 | 28 | 45 |
| Доля одновид. родов, % | 81,5 | 76,6 | 78,6 | 75,6 | 80,7 | 79,1 | 81,5 | 76,9 | 73,6 |
| Доля одновид. сем., % | 58,1 | 50 | 42,9 | 48,9 | 48,4 | 44 | 53,1 | 32,1 | 48,9 |
| ОПП мохового яруса, % | 15 | <1 | 5 | 5 | 1 | 3 | 1 | 50 | 75 |
| Сред.число видов в асс. | 38 | 21 | 47 | 63 | 53 | 42 | 31 | 28 | 67 |
| Верхоплодные мхи | 21 | 22 | 38 | 46 | 35 | 28 | 28 | 37 | 63 |
| Бокоплодные мхи | 40 | 33 | 57 | 52 | 40 | 30 | 33 | 30 | 52 |
| Печеночники | 10 | 9 | 9 | 23 | 5 | 5 | 10 | 7 | 23 |
| Однодомные | 31 | 27 | 41 | 44 | 33 | 22 | 30 | 27 | 46 |
| Двудомные | 38 | 37 | 61 | 73 | 47 | 40 | 40 | 47 | 85 |
| Многодомные | 2 | 0 | 2 | 4 | 0 | 1 | 1 | 0 | 7 |
| Редкие в РБ виды | 3 | 5 | 14 | 10 | 10 | 1 | 2 | 1 | 13 |
| Brachytheciaceae | 11 | 10 | 16 | 11 | 10 | 9 | 10 | 7 | 12 |
| Dicranaceae | 3 | 4 | 11 | 9 | 10 | 9 | 8 | 10 | 14 |
| Amblystegiaceae | 7 | 4 | 8 | 6 | 5 | 4 | 5 | 4 | 7 |
| Mniaceae | 6 | 1 | 6 | 10 | 1 | 2 | 2 | 3 | 11 |
| Scapaniaceae s.l. | 2 | 2 | 3 | 9 | 1 | 1 | 2 | 2 | 10 |
| Grimmiaceae | - | 2 | 2 | 2 | 4 | 2 | 2 | 2 | 2 |
| Polytrichiaceae | 2 | 1 | 3 | 6 | 2 | 3 | 4 | 4 | 9 |
| Pottiaceae | 1 | 5 | 2 | 3 | 6 | 2 | 2 | 2 | 2 |
| Bryaceae | 3 | 4 | 3 | 2 | 3 | 4 | 2 | 5 | 4 |
| Plagiotheciaceae | 4 | 2 | 2 | 3 | 2 | 1 | 2 | 2 | 6 |
| Pylaisiaceae | 5 | 4 | 7 | 6 | 5 | 5 | 5 | 5 | 6 |
| Sphagnaceae | - | - | - | 2 | - | - | - | - | 6 |
| Hylocomiaceae | 2 | 2 | 5 | 6 | 3 | 3 | 3 | 3 | 6 |
Примечание. Номера союзов: 1 – Alnion incanae, 2 – Lathyro-Quercion, 3 – Aconito-Tilion, 4 – Aconito-Piceion, 5 – Caragano-Pinion, 6 – Veronico-Pinion, 7 – Trollio-Pinion, 8 – Dicrano-Pinion, 9 – Piceion excelsae. Сокращения: ОПП – общее проективное покрытие, сред. – среднее, асс. ассоциация, одновид. - одновидовой, сем. – семейство.
Использование коэффициента Съеренсена-Чекановского для сравнения бриоценофлор союзов лесной растительности РБ показало, что они не отличаются высоким сходством (таблица 3). Наиболее близки были группы бриоценофлор: 1. лиственных лесов (союзы Lathyro-Quercion, Alnion incanae и Aconito-Tilion) (K 0,53-0,61), 2. темнохвойно-широколиственных лесов и темнохвойных лесов (союзы Piceon excelsae и Aconito-Piceon) (K 0,67), 3. сосновых лесов разных типов (K 0,62-0,68).
В обследованных лесных сообществах при повышении доли темнохвойных пород в древостое возрастают видовое богатство мохообразных, доля верхоплодных листостебельных мхов, представителей семейств Dicranaceae, Mniaceae, Polytrichaceae, Hylocomiaceae, а также печеночников.
Таблица 3 Полуматрица значений коэффициента Съеренсена-Чекановского значения коэффициента *100) для бриоценофлор лесных союзов РБ
|
AI |
L-Q |
A-T |
A-P |
C-P |
V-P |
T-P |
D-P |
PE |
|
|
AI |
53 | 61 | 45 | 37 | 50 | 40 | 34 | 35 | |
|
LQ |
57 | 39 | 51 | 50 | 45 | 37 | 30 | ||
|
AT |
62 | 55 | 58 | 43 | 42 | 44 | |||
|
AP |
51 | 56 | 46 | 48 | 67 | ||||
|
CP |
64 | 63 | 62 | 45 | |||||
|
VP |
61 | 64 | 48 | ||||||
|
TP |
68 | 49 | |||||||
|
DP |
58 | ||||||||
|
PE |
Примечание. AI – Alnion incanae, L-Q – Lathyro-Quercion, A-T – Aconito-Tilion, A-P – Aconito-Piceion, C-P – Caragano-Pinion, V-P – Veronico-Pinion, T-P – Trollio-Pinion, D-P – Dicrano-Pinion, PE – Piceion excelsae.
Постоянство видов и видовое богатство бриоценофлор
Невысокий уровень сходства бриоценофлор союзов можно объяснить высокой долей видов, имеющих низкое постоянство в сообществах: около 25 % видов было встречено от 1 до 3 раз, а доля видов, встреченных менее, чем в 5 % описаний, составляла от 68 до 85% в разных союзах (рисунок 1). Максимальная разница в пропорциях редко встреченных и высококонстантных видов характерна для сообществ, которые характеризуются высоким видовым богатством мохообразных и либо имеют большие ценоареалы, в пределах которых представлено большое разнообразие местообитаний (сообщества широколиственных лесов ассоциаций Brachypodio pinnati-Tilietum cordatae и Stachyo sylvaticae-Tilietum cordatae), либо встречаются в виде изолированных фрагментов в окружении других типов растительности (высокотравные горные уремники ассоциации Crepido sibiricae-Alnetum incanae). Наибольшее разнообразие видов мохообразных отмечено в темнохвойных зеленомошных лесах союза Piceion excelsae (138 видов) и темнохвойно-широколиственных лесах союза Aconito-Piceion (121).
Самая бедная и малоспецифичная бриофлора характерна для мезоксерофитных дубняков союза Lathyro-Quercion и березово-сосновых травяных лесов союза Veronico-Pinion (по 63 вида). Необходимо отметить, что высокое видовое богатство в пределах класса Querco-Fagetea можно объяснить включением в его состав темнохвойно-широколиственных и сосново-широколиственных лесов. В типичных сообществах широколиственных лесов и пойменных уремниках обнаружено всего 117 видов, что значительно ниже, чем в бореальных лесах класса Vaccinio-Piceetea.
Рисунок 1 – Постоянство видов мохообразных в сообществах лесных союзов РБ

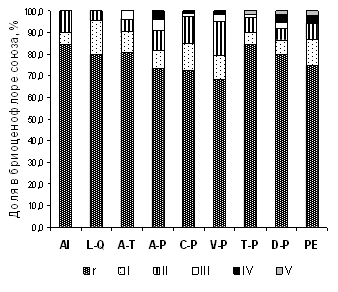
На уровне ассоциаций максимальное разнообразие мохообразных отмечено в сообществах союзов Piceion excelsae (асс. Asaro europaei-Piceetum obovatae – 95 видов, Linnaeo borealis-Piceetum abietis – 70 видов), Aconito-Piceion (асс. Cerastio pauciflori-Piceetum obovatae – 80 видов, Chrysosplenio alternifolii-Piceetum obovatae – 78 видов, Violo collinae-Piceetum obovatae – 76 видов), Caragano-Pinion (асс. Ceraso fruticis-Pinetum sylvestris – 78 видов), Alnion incanae (асс. Crepido sibiricae-Alnetum incanae – 67 видов) и Aconito-Tilion (асс. Tilio cordatae-Pinetum sylvestris – 65 видов). Эти леса характеризуются слабой степенью нарушенности и высоким разнообразием представителей всех субстратных групп эпифитов старых широколиственных деревьев, эпиксилов, эпилитов карбонатосодержащих пород и напочвенных видов. Сообщества отмечены на склонах в долинах горных рек (Большой и Малый Инзер, Зилим, Нугуш, Уфа, Юрюзань, Урюк, Тюльма, верховья Белой и др.). В литературе отмечается, что в периоды регрессивных фаз и континентализации климата в плейстоцене плакоры подвергались остепнению, а лесная растительность сохранялась в значительно глубже (по сравнению с настоящим временем) врезанных речных долинах [Восточноевропейские леса…, 2003]. Вероятно, именно этим явлением можно объяснить встречаемость в водоохранно-защитных лесах горных рек редких для Урала неморальных реликтовых видов с дизъюнктивным ареалом: Eurhynchium angustirete, Brachythecium geheebii, Anomodon rugelii, Frullania bolanderi, Pylaisia selwynii, Entodon schleicheri, Metzgeria furcata, Lejeunea cavifolia и др.
Распределение видов по типам субстратов
При анализе субстратной приуроченности бриофитов необходимо принять во внимание, что многие виды способны произрастать на разных типах субстратов [Barkman, 1958]. На основаниях стволов, где кора начинает разрушаться и приобретает некоторые свойства гнилой древесины, были представлены практически все виды эпифитного комплекса и две трети эпиксильного. В составе эпиксильных сообществ третью часть составляли виды, обычные в напочвенном покрове [Баишева, Игнатова, 1998].
В обследованных лесах обнаружено 32 вида мохообразных, растущих на стволах живых деревьев и гнилой древесине начальных стадий разрушения; 38 – на камнях и гнилой древесине; 19 – на почве и гнилой древесине; 61 – преимущественно на почве; 57 – на камнях и покрывающем их слое мелкозема; 5 – на камнях и стволах живых деревьев. К облигатным эпифитам в районе исследования можно отнести Leucodon sciuroides, Ortotrichum obtusifolium, Orthotrichum speciosum, к облигатным эпиксилам – Blepharostoma trichophyllum, Lepidozia reptans, Tetraphis pellucida и др. Сквозное распространение в сообществах всех типов обследованных лесов имеют Stereodon pallescens, Sanionia uncinata, Dicranum montanum, Sciuro-hypnum reflexum, Ptilidium pulcherrimum, Plagiomnium cuspidatum, Brachythecium salebrosum, Callicladium haldanianum, Pseudoleskeella nervosa, Pylaisia polyantha, Pohlia nutans и некоторые др. виды, растущие на стволах живых деревьев и гнилой древесине.
Типичные виды бореальных лесов Pleurozium schreberi, Hylocomium splendens, Dicranum polysetum, Dicranum scoparium, Rhytidiadelphus triquetrus практически отсутствуют в сообществах союза Lathyro-Quercion, изредка встречаются в горных уремниках и имеют сквозное распространение в остальных типах леса. В бореальных зеленомошных лесах они растут на подстилке, наползая на гнилую древесину и камни, в смешанных высокотравных лесах они встречаются преимущественно на гнилой древесине, избегая конкуренции с сосудистыми растениями.
Данные рисунка 2 показывают, что для бриоценофлор южно-уральских лесных сообществ характерно высокое участие эпилитов (от 11 до 29 % в разных союзах). Преобладание видов эпифитно-эпиксильного комплекса над напочвенными видами отмечено для сообществ союзов Lathyro-Quercion, Caragano-Pinion и Veronico-Pinion, в которых развитию напочвенных мохообразных препятствуют засушливые условия местообитания и, в ряде случаев, высокая сомкнутость травяного яруса. Повышенное разнообразие эпигейных видов характерно для ельников-зеленомошников (союз Piceion excelsae).
Рисунок 2 – Спектр субстратной приуроченности бриофитов лесных союзов РБ
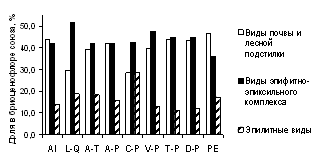
Примечание. AI – Alnion incanae, L-Q – Lathyro-Quercion, A-T – Aconito-Tilion, A-P – Aconito-Piceion, C-P – Caragano-Pinion, V-P – Veronico-Pinion, T-P – Trollio-Pinion, D-P – Dicrano-Pinion, PE – Piceion excelsae.
Половые типы мохообразных лесных сообществ РБ
Среди мохообразных двудомность распространена шире, чем в других отделах растительного мира [Бойко, 1999]. В мире среди листостебельных мхов двудомными являются более половины видов, среди печеночников – две трети [Longton, Schuster, 1983; Crum, 2001]. В России на долю двудомных листостебельных мхов приходится примерно 54%, причем в различных регионах соотношение одно- и двудомных видов мхов примерно одинаково [Игнатов и др., 2009].
На уровне союзов, для всех типов обследованных лесных сообществ РБ характерно численное превосходство двудомных видов мохообразных над однодомными (рисунок 3).
Установлено, что виды из разных субстратных групп имеют существенные различия в спектрах половых типов видов. В союзах доля однодомных видов варьировала среди эпифитов от 43 до 63 %, среди эпиксилов – от 59 до 87 %, среди эпигейных видов – от 14 до 32 %, среди эпилитных – от 17 до 41 %. Таким образом, однодомные виды преобладали среди видов эпифитно-эпиксильного комплекса, а двудомные – среди напочвенных и эпилитных мохообразных.
Рисунок 3 – Соотношение видов с разными половыми типами в лесных союзах РБ

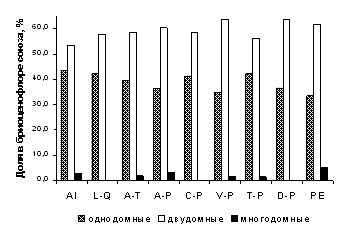
Только для 7 из 42 ассоциаций лесной растительности была выявлена высокая (более 50 %) доля однодомных видов. Такая особенность характерна для сообществ, встречающихся узкими полосами вдоль рек (пойменные ольхово-черемуховые уремники, долинные сосново-березовые леса с периодическим подтоплением), дубняков лесостепной зоны и некоторых типов светлохвойно-широколиственных лесов, которые встречаются небольшими фрагментами среди массивов широколиственных лесов или степей, часто – на крутых склонах. Для всех этих сообществ характерна малоспецифичная бриофлора, представленная, главным образом, ксеромезофитными эпиризными и эпиксильными видами со стратегией колонистов. Развитию двудомных напочвенных мхов в этих сообществах препятствуют такие факторы, как сомкнутый травяной ярус, опад деревьев широколиственных пород, высокий уровень аэрации (в лесостепных сообществах) или поемный режим (в уремниках).
Лесные сообщества РБ часто имеют сложную структуру, встречаются в условиях горного рельефа, в местообитаниях с многочисленными выходами камней. Поэтому использование такого критерия, как соотношение долей однодомных и двудомных видов мохообразных (без учета их субстратной приуроченности) может оказаться недостаточно информативным для того, чтобы делать выводы об экологическом режиме сообществ.
Автор полагает, что для этой цели более оправдано учитывать виды с разными способами размножения. Исследования популяционной биологии мохообразных показали, что расселение за счет спор жизненно необходимо для возникновения новых популяций (при заселении свежих, недавно возникших субстратов, восстановительных сукцессиях, инвазиях в новые растительные сообщества). Вегетативное размножение часто играет ведущую роль для выживания и разрастания популяции, так как оно более эффективно в стабильных условиях, когда происходит клонирование набора генов тех растений, которые оказались хорошо приспособлены к данным конкретным условиям существования [Kimmerer, 2003; Longton, 2006].
Рисунок 4 – Соотношение видов с разными типами размножения среди мохообразных разных субстратных групп
|
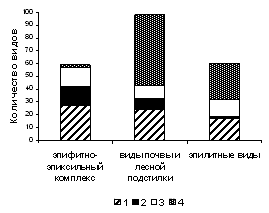
На рисунке 4 показано, что виды групп 1 и 2 преобладают среди эпифитов и эпиксилов. В то же время, почва лесных сообществ и, особенно, каменистые субстраты представляют собой стабильные условия существования, к которым хорошо приспособлены виды мохообразных с преимущественно вегетативным способом размножения, а также виды, не имеющие специализированных органов вегетативного размножения и редкими (или отсутствующим в регионе) спороношением (группы 3 и 4).
Повышенные пропорции видов с активным спороношением отмечены в сообществах союзов Alnion incanae, Lathyro-Quercion, Trollio-Pinion и Dicrano-Pinion. Виды, не имеющие специализированных органов вегетативного размножения и редкое (или отсутствующее в регионе) спороношение, в лучшей степени представлены в сообществах союза Piceion excelsae, в меньшей степени – в сообществах союзов Lathyro-Quercion и Aconito-Tilion.
На уровне бриоценофлор ассоциаций был проведен подсчет соотношения числа видов, часто образующих спорогоны, к числу видов с редким (или отсутствующим в регионе) спороношением. Значение коэффициента меньше единицы оказалось у ассоциаций старовозрастных лесов (Anemonastro biarmiensis-Laricetum sukaczewii, Asaro europaei-Piceetum obovatae, Zigadeno sibirici-Pinetum sylvestris, Cerastio pauciflori-Piceetum obovatae, Linnaeo borealis-Piceetum abietis). Значение коэффициента свыше 1,5 оказалось у лесов, которые либо представлены изолированными участками в поймах (Alnetum incanae), лесостепной зоне (Brachypodio pinnati-Quercetum roboris), среди сельхозугодий (Lasero trilobi-Quercetum roboris), либо имеют высокую интенсивность естественных нарушений (Carici arnellii-Pinetum sylvestris, Galio odorati-Pinetum sylvestris).
Таким образом, этот коэффициент может использоваться для оценки стабильности экологических условий в лесном сообществе и косвенно позволяет судить о длительности ненарушенного существования местообитаний. Такие данные, наряду с анализом флористического состава, позволяют выявить наиболее ценные для охраны участки лесов и могут быть использованы для мониторинга за их состоянием.
Жизненные стратегии мохообразных лесных сообществ РБ
Интегральной характеристикой видов могут служить их эколого-фитоценотические стратегии, т.е. способ выживания в условиях разных экотопов, сообществ и режимов экзогенных нарушающих воздействий. Знание типов стратегий видов и учет соотношения видов с разными стратегиями является необходимым условием для понимания и построения моделей фитоценозов [Раменский, 1938; Grime, 1979; Миркин, Наумова, 1998]. Автором проведен анализ жизненных стратегий бриофитов, выделенных в соответствии с системой H. During [1979, 1991], основой которой является реакция мхов на изменения окружающей среды, выражающиеся в продолжительности существования и степени вероятности появления местообитаний, пригодных для бриофитов.
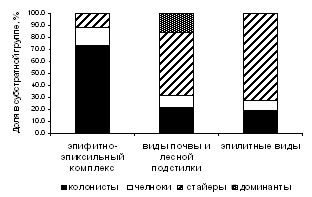
Анализ спектров стратегий у видов разных субстратных групп (рисунок 5) показал, что среди эпифитов и эпиксилов преобладали виды со стратегией колонистов и челноков, среди эпигейных и эпилитных видов – многолетние стайеры. Вероятно, это связано с тем, что период существования стволов деревьев и гнилой древесины ограничен несколькими годами или десятилетиями, поэтому стратегия выживания эпифитных и эпиксильных видов направлена на активное расселение. В то же время, доля колонистов и челноков среди напочвенных видов и эпилитных лесных видов невысока, так как продолжительность существования их субстратов намного дольше.
Рисунок 5 – Спектры жизненных стратегий мохообразных разных субстратных групп
Повышенная доля стайеров отмечена в сообществах союзов Piceion excelsae, Caragano-Pinion, Veronico-Pinion и Aconito-Piceion, характеризующихся либо развитым ярусом напочвенных мхов, либо наличием многочисленных скальных выходов (рисунок 6).
На уровне ассоциаций пропорция стайеров, превышающая 45 % бриоценофлор, отмечена во всех старовозрастных сообществах ассоциаций Anemonastro biarmiensis-Laricetum sukaczewii, Bistorto majoris-Piceetum obovatae, Zigadeno sibirici-Pinetum sylvestris, Cerastio pauciflori-Piceetum obovatae, Linnaeo borealis-Piceetum abietis, а также в 3 ассоциациях, характеризующихся бедным бриокомпонентом и слабым развитием эпифитно-эпиксильного комплекса (Violo rupestris-Pinetum, Carici macrourae-Quercetum и Myosotido sylvaticae-Pinetum).
Рисунок 6 – Спектры жизненных стратегий мохообразных в бриоценофлорах лесных союзов РБ

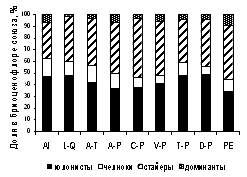
Высокая (более 70 % от бриоценофлоры ассоциации) доля колонистов и челноков, в основном, характерна для сообществ, представленных фрагментами в лесостепи или среди сельхозугодий. Это сообщества союзов Lathyro-Quercion, Alnion incanae и ассоциация Brachypodio-Tilietum, объединяющая ксеромезофитные широколиственные леса, встречающиеся на склонах в лесостепной зоне.
Проведенные исследования показали, что для оценки экологического режима местообитаний, длительности стабильного существования и степени фрагментированности лесных сообществ необходимо полное выявление лесных бриоценофлор, анализ спектров жизненных стратегий бриофитов и подсчет соотношения видового богатства групп видов с разной активностью и способами размножения. Учет соотношения только половых типов мохообразных может оказаться недостаточно информативным. Во всех случаях необходимо учитывать субстратную приуроченность видов, т.к. для лесных сообществ с неразвитым ярусом напочвенных мхов и отсутствием выходов камней, использование этих методов анализа может иметь существенные ограничения.
Использование мохообразных в качестве индикаторов лесов высокой природоохранной ценности
Биоразнообразие лесных экосистем крайне уязвимо вследствие интенсивного вовлечения лесов в хозяйственную деятельность, загрязнения окружающей среды, рекреационной нагрузки и прочих изменений растительного покрова, вызванных деятельностью человека. В связи с этим, особую актуальность приобретают исследования, направленные на выявление лесов, представляющих высокую природоохранную ценность [Выявление…, 2009].
В отношении мохообразных, распространение которых, в основном, определяется наличием и качеством подходящих для них субстратов и местообитаний [van Zanten, Pocs, 1981; Miles, Longton, 1992 и др.], выявление таких участков имеет особое значение. В старых ненарушенных лесах часто находятся микро- и мезоместообитания, которые очень долгое время были доступны для непрерывной колонизации бриофитами. При рассмотрении данной проблемы можно выделить два аспекта – выявление индикаторов старовозрастных лесов и индикаторов участков с длительным ненарушенным существованием, так называемых зон экологической связности местообитаний – «Ecological Continuity» [Norden, Appelqvist, 2001]. Зачастую эти два понятия смешиваются, что не вполне правомерно, учитывая различия в жизненных стратегиях мохообразных и длительность ненарушенного существования леса, которая принимается в расчет.
В большей степени разработаны вопросы, касающиеся использования мохообразных в качестве биоиндикаторов старовозрастных лесов. Исследования показали, что существует положительная взаимосвязь между обилием видов, высоким богатством бриофлоры и крупными участками старовозрастных лесов, которые длительное время не страдали от таких крупномасштабных нарушений, как, например, пожары или рубки [Edwards, 1986; Vellak, Paal, 1999 и др.]. Чаще всего в качестве индикаторов старовозрастных лесов используют печеночники, особенно редкие эпиксильные виды, распространение которых связано с наличием гнилой древесины различных степеней разрушения и размером самого участка старовозрастных лесов [Gustafsson, Hallingbäck 1988; Newmaster et al. 2003 и др.].
Учитывая древность мохообразных, а также высокую долю видов, имеющих ограниченные способности к расселению, для охраны бриофитов исключительно важное значение приобретает выявление местообитаний, на которых нарушений экологического режима (особенно колебаний влажности и освещенности) не было в течение очень длительного времени.
Не все участки старовозрастных лесов подходят для этих целей. Для того, чтобы отнести лес к категории старовозрастного, необходимо, чтобы возраст древостоя был более 120-260 лет (в зависимости от породы дерева и региона) [Выявление…, 2009]. В то же время, для выживания реликтовых популяций мохообразных, часто имеющих дизъюнктивный ареал, ограниченные способности к расселению и высокие требования к стабильности экологического режима местообитаний, необходимо, чтобы они существовали в так называемых девственных лесах («ancient forest», «primeval forest»). Речь идет о естественных лесах, не испытавших заметного антропогенного воздействия и изменяющихся на протяжении многих поколений лесообразующих древесных пород только вследствие природных процессов [Выявление…, 2009]. В настоящее время эти леса сохранились лишь в виде небольших фрагментов в труднодоступных для человека местах в окружении более или менее антропогенно трансформированной лесной растительности [Trass et al., 1999]. Также следует отметить, что некоторые леса (например, сосняки), для которых характерны периодические возникающие в силу естественных причин пожары, для выживания реликтовых бриофитов не подходят.
Анализ жизненных стратегий мохообразных, считающихся индикаторами старовозрастных лесов, показал, что среди них представлены виды как с высокой, так и с низкой активностью размножения. Так, например, стратегию колонистов имеют Homalia trichomanoides, Herzogiella seligerii, Entodon schleicheri, стратегию челноков – Neckera complanata, Frullania bolanderi, Lejeunea cavifolia. Есть виды интенсивно размножающиеся вегетативным способом (Dicranum flagellare, Dicranum viride, Orthocaulis attenuatus и др.). В основном, перечисленные таксоны относятся к эпифитно-эпиксильному комплексу и способны расселяться из близлежащих участков леса, но только при наличии подходящих субстратов (чаще всего они отмечены на стволах старых широколиственных деревьев или на сильно перегнившей древесине). Эти виды могут считаться индикаторами старовозрастных лесов, но не подходят для выявления зон экологической связности местообитаний.
Виды со стратегией стайеров, не имеющие специализированных органов вегетативного размножения, в основном растут на почве или, в случае горных лесов, на камнях. По всей видимости, именно они могут служить индикаторами как старовозрастных, так и девственных лесов. На территории РБ они в основном встречаются на крутых склонах по берегам горных рек Уфимского плато и центрально-возвышенной части Южного Урала.
Виды, чувствительные к различным видам антропогенного воздействия, относят к категории гемерофобных [Trass et al., 1999]. Ниже приведены списки видов мохообразных, которые, по мнению автора, на Южном Урале являются гемерофобными и могут служить индикаторами участков биологически ценных лесов РБ.
Индикаторы зон длительного ненарушенного существования местообитаний: Plagiomnium confertidens, Dicranum drummondii, Bryhnia scabrida, Brachythecium geheebii, Entodon concinnus, Anomodon rugelii, Eurhynchium angustirete, Orthothecium intricatum, Myurella sibirica, Campylidium calcareum. Практические все эти виды имеют стратегию многолетних стайеров, низкую активность размножения, многие – дизъюнктивный ареал.
Индикаторы старовозрастных лесов: Neckera pennata, Anomodon longifolius, Anomodon viticulosus, Anomodon attenuatus, Dicranum viride, Leucodon sciuroides, Homalia trichomanoides, Haplocladium microphyllum, Orthotrichum affine, Plagiothecium nemorale, Orthotrichum pallens, Pseudobryum cinclidioides, Polytrichastrum formosum, Polytrichastrum pallidisetum, Dicranum fragilifolium, Dicranum flagellare, Mnium spinosum, Polytrichastrum longisetum, Dicranum bergeri, Sphenolobus minutus, Hylocomiastrum umbratum, Hylocomiastrum pyrenaicum, Rhytidiadelphus subpinnatus, Barbilophozia lycopodioides, Lepidozia reptans, Barbilophozia barbata, Frullania bolanderi, Orthocaulis attenuatus, Barbilophozia hatcheri, Lophozia longidens, Pylaisia selwynii. Большая часть этих видов имеют стратегию многолетних стайеров и доминантов, но представлены также колонисты и челноки.
Индикаторы уникальных местообитаний со стабильно высоким уровнем влажности: Taxyphyllum wissgrilli, Rhynchostegium arcticum, Tritomaria quinquedentata, Lejeunea cavifolia, Jamesoniella autumnalis, Metzgeria furcata, Calypogeia integristipula, Tritomaria exsectiformis. В основном, это многолетние стайеры и печеночники со стратегией челноков.
Анализ взаимосвязи яруса напочвенных мхов с другими компонентами лесных сообществ (на примере лесов Уфимского плато)
Леса Уфимского плато (УП) представляют уникальный по разнообразию комплекс сообществ, объединяющий неморальнотравные широколиственно-темнохвойные, темнохвойные и светлохвойно-широколиственные леса, а также типичные бореальные зеленомошные светлохвойные и темнохвойные леса на слаборазвитых и мерзлотных перегнойно-карбонатных почвах [Кулагин, 1978; Мартьянов и др., 2002; Жигунова, 2006]. Данные по 230 полным геоботаническим описаниям лесной растительности показали, что в лесах УП при разреженном древостое с сомкнутостью менее 75% покрытие напочвенных мхов может сильно варьировать от 1 до 95 % (в среднем 50 %). При сомкнутости древостоя от 75% и выше проективное покрытие напочвенных мхов не превышало 35 % (в среднем 3 %). При анализе всего массива данных была выявлена отрицательная корреляция между проективным покрытием напочвенных мхов и такими показателями, как сомкнутость древесного (r= -0,64*) и травяного (r= -0,58*) ярусов, средняя высота травяного яруса (r = -0,56*), а также роль в древостое Acer platanoides, Quercus robur, Tilia cordata и Ulmus glabra. Сходные результаты были получены и при использовании метода прямого градиентного анализа (CCA). Результаты более детального анализа корреляционных зависимостей представлены в таблице 4.
Таблица 4 Коэффициенты корреляции проективного покрытия напочвенных лесных мхов с другими параметрами лесных сообществ УП
|
Параметры лесных сообществ |
Группы сообществ |
|
|
А |
В |
|
| ОПП деревьев хвойных пород | 0,61* | 0,30* |
| ОПП деревьев лиственных пород | –0,67* | NS |
| ОПП травяного яруса | –0,67* | NS |
| ОПП неморального высокотравья | –0,55* | –0,28* |
| ОПП злаков и осок | NS | NS |
| ОПП бореального мелкотравья | 0,59* | 0,35* |
| ОПП лугового и лесного разнотравья | NS | NS |
| Средняя высота древостоя | NS | NS |
| Крутизна склона | 0,46* | 0,25** |
| Число видов бриофитов на площадке | NS | 0,18*** |
| Число видов сосудистых растений на площадке | NS | NS |
Примечание. ОПП – общее проективное покрытие; А сообщества с разреженным древостоем (ОПП древесного яруса <75%), В сообщества с сомкнутым древостоем (ОПП древесного яруса >75 %). Уровни значимости коэффициентов корреляции: *р<0,001, **р<0,01, ***р<0,05, NS – не значим.
В группе А представлены практически все описанные на УП зеленомошные ельники и сосняки, а также небольшая часть неморальнотравных еловых и темнохвойно-широколиственных лесов. В группу B вошли все неморальнотравные сосновые и темнохвойно-широколиственные леса, а также большинство неморальнотравных ельников. Показано, что при разреженном древостое повышение обилия напочвенных мхов происходит одновременно с увеличением доли хвойных и бореального мелкотравья на площадках, а также при возрастании крутизны склона и, соответственно, лучшей освещенности напочвенного покрова. При высоком уровне затенения со стороны деревьев (более 75%), коррелятивные связи между проективным покрытием напочвенных мхов и другими компонентами фитоценоза, по всей видимости, ослабляются.
Применение метода непрямого градиентного анализа (DCA) для всего массива описаний показало, что основными факторами, определяющими различия в распределении видов напочвенных мхов в обследованных сообществах, по всей видимости, являются, во-первых, комплексный градиент, отражающий богатство и развитость почв, во-вторых, освещенность напочвенного покрова на площадках. При этом влияние первого фактора, вероятнее всего, осуществляется опосредованно через возрастание проективного покрытия трав и ухудшение освещенности напочвенных мхов.
Глава 6. Синтаксономия бриосообществ, распространенных в лесах РБ
Автором была разработана эколого-флористическая классификация эпифитных, эпиксильных и прибрежно-водных бриосообществ, встречающихся в лесной и лесостепной зонах РБ. Напочвенные лесные мхи описывались в качестве отдельного яруса в составе сообществ сосудистых растений, отдельно их группировки не рассматривались. В главе приведена подробная характеристика каждого синтаксона с указанием общего распространения, особенностей экологии и результатов сравнения с аналогами, описанными в Западной и Центральной Европе. Бриосинтаксономия РБ включает 16 ассоциаций, 9 союзов, 6 порядков и 4 класса. 5 ассоциаций выделены впервые.
Продромус синтаксонов моховой растительности РБ
КЛАСС PLATHYHYPNIDIO – FONTINALIETEA ANTIPYRETICAE Philippi 1956
ПОРЯДОК HYGROHYPNETALIA Krajina 1933
СОЮЗ Racomitrion acicularis v. Krusenstjerna 1945
Асс. Scapanietum undulatae Schwickerath 1944
ПОРЯДОК LEPTODICTYETALIA RIPARII Philippi 1956
СОЮЗ Brachythecion rivularis Hertel 1974
Асс. Brachythecio rivularis - Hygrohypnetum luridi Philippi 1965
Асс. Cratoneuretum filicini Poelt 1954
СОЮЗ Fontinalion antipyreticae W. Koch 1936
Асс. Fontinalietum antipyreticae Kaiser ex Frahm 1971
КЛАСС FRULLANIO DILATATAE – LEUCODONTETEA SCIUROIDIS Mohan 1978
ПОРЯДОК ORTHOTRICHETALIA Hadač in Klika & Hadač 1944
СОЮЗ Leskeion polycarpae Barkman 1958
Асс. Syntrichio latifoliae - Leskeetum polycarpae v. Hübschmann 1952
Асс. Brachythecio salebrosi- Amblystegietum serpentis Baisheva & al. 1994
субасс. B.s.-A.s. typicum Baisheva & al. 1994
субасс. B.s.-A.s. plagiomnietosum cuspidati Baisheva 1995
СОЮЗ Syntrichion laevipilae Ochsner 1928
Асс. Pylaisielleto polyanthae - Leskeelletum nervosae Baisheva & al. 1994
Асс. Pylaisietum polyanthae Felföldy 1941
Асс. Orthotrichetum speciosi Barkman 1958
субасс. O.s. orthotrichetosum obtusifolii Baisheva 1995
Асс. Orthotrichetum pallentis Ochsner 1928
КЛАСС CLADONIO DIGITATAE – LEPIDOZIETEA REPTANTIS Ježek & Vondráček 1962
ПОРЯДОК CLADONIO DIGITATAE LEPIDOZIETALIA REPTANTIS Ježek & Vondráček 1962
СОЮЗ Nowellion curvifoliae Philippi 1965
Асс. Brachythecietum reflexi Baisheva & al. 1994
Асс. Plagiothecio laeti-Pohlietum nutantis Baisheva & al. 1994
СОЮЗ Tetraphidion pellucidae v. Krusenstjerna 1945
Сообщество Tetraphis pellucida
Сообщество Pohlia nutans - Plagiothecium denticulatum
ПОРЯДОК DICRANETALIA SCOPARII Barkman 1958
СОЮЗ Dicrano scoparii – Hypnion filiformis Barkman 1958
Асс. Ptilidio pulcherrimi-Hypnetum pallescentis Barkman ex Wilmanns 1962
субасс.P.p.-H.p. typicum Barkman ex Wilmanns 1962
субасс. P.p.-H.p. callicladietosum haldaniani Baisheva 1995
Асс. Platygyrietum repentis Le Blanc ex Marstaller 1986
Асс. Orthodicrano montanii-Plagiothecietum laeti Baisheva et al.1994;
КЛАСС Hylocomietea splendentis Marstaller 1992
ПОРЯДОК HYLOCOMIETALIA SPLENDENTIS Gillet ex Vadam 1990
СОЮЗ Pleurozion schreberii v. Krusenstjerna 1945
Асс. Pleurozietum schreberi Wiśniewski 1930
Отмечается, что все прибрежно-водные бриосообщества РБ отнесены к ассоциациям, описанным в Европе, небольшие различия с которыми отражены на уровне вариантов. Эпифитные и эпиксильные бриосообщества РБ, напротив, довольно сильно различаются с аналогами из Центральной Европы, в первую очередь, за счет сильно обедненного флористического состава. Континентальный климат Южного Урала неблагоприятен для эпифитных мхов, развитие которых ограничивается сухостью воздуха летом и опасностью вымерзания зимой. Обычными местообитаниями эпифитов являются основания и нижние части стволов, защищенные травяным ярусом. На высоте более 1.5 метров эпифитные группировки были отмечены только в специфических условиях: в теневых влажных горных и пойменных лесах; на изогнутых, горизонтально наклоненных стволах, защищенных зимой снежным покровом; на стволах деревьев с гигроскопичной корой, впитывающей влагу осадков и создающих благоприятный водный режим (Tilia cordata, Populus tremula).
При обследовании эпифитных мохообразных РБ была подтверждена общая закономерность - отсутствие выраженной приуроченности видов мхов к определенным породам деревьев [Бойко, 1978; Улычна, 1989; Piippo, 1982 и др.]. Сборы проводились на коре 16 видов деревьев. В зоне средних частей стволов наибольшее количество видов было отмечено на коре Tilia cordata (16 видов), Betula pubescens (9), Populus tremula (6), Acer platanoides (5), Quercus robur (5). Несколько иная картина наблюдалась в отношении видов, поселяющихся на основаниях стволов: наиболее благоприятным субстратом является кора Betula pendula (43 вида), Tilia cordata (38), Abies sibirica (29), Alnus incana (25), Betula pubescens (21). Самый широкий спектр базовых пород выявлен у Pylaisia polyantha (вид отмечен на 14 породах), Orthotrichum speciosum (12), Brachythecium salebrosum (11), Amblystegium serpens, Sciuro-hypnum reflexum (10), Plagiomnium cuspidatum, Sanionia uncinata, Leskea polycarpa (9).
На гнилой древесине выявлено 69 видов, из которых только 18 видов не были отмечены на коре живых деревьев. В основном это виды, приуроченные к сильно разрушенной древесине (Tetraphis pellucida, Leptobryum pyriforme, Cephalozia lunulifolia), а также встречающиеся на почве (Polytrichum juniperinum, Abietinella abietina и др.) .
Глава 7. Анализ бриофлоры лесной, лесостепной зон РБ и ИХ ГОРНЫХ АНАЛОГОВ
Видовое разнообразие и таксономический анализ
В главе 5 были рассмотрены бриоценофлоры коренных зональных лесов РБ и их горных аналогов, выявленные с помощью методов учета на пробных площадях, закладываемых на типичных лесных участках. Одновременно проводился сбор мохообразных и в других типах местообитаний. Материалы этих исследований позволили выявить флору мохообразных лесной и лесостепной зон РБ, а также их горных аналогов. Конспект флоры представлен в Приложении, распределение видов дано по районам в соответствии с системой районирования А.А. Мулдашева [Реестр, 2006], с некоторыми дополнениями по работе П.Л. Горчаковского [1988].
В районе исследования обнаружено 365 видов листостебельных мхов, относящихся к 141 родам и 45 семействам, и 91 вид печеночников, относящихся к 44 родам и 28 семействам. Основные показатели флоры представлены на таблице 5.
Ведущие семейства флоры листостебельных мхов: Grimmiaceae (31 вид), Pottiaceae (31), Brachytheciaceae (27), Sphagnaceae (27), Amblystegiaceae (26), Dicranaceae (23), Bryaceae (19), Mniaceae (16), Mielichhoferiaceae (13), Rhabdoweisiaceae (13), Polytrichaceae (12), Pylaisiaceae (12), ведущие роды: Sphagnum (27 видов), Dicranum (17), Bryum (17), Grimmia (14), Pohlia (13), Schistidium (12), Brachythecium (10), Orthotrichum (8), Plagiomnium (7), Fissidens (7), Sciuro-hypnum (6), Tortula (6).
Таблица 5 Характеристика бриофлоры лесной и лесостепной зон РБ
| Показатели флоры | листостебельные мхи | печеночники | ||||
| Район исследования | ||||||
| 1 | 2 | всего | 1 | 2 | всего | |
| Общее число видов | 191 | 332 | 365 | 37 | 83 | 91 |
| Общее число родов | 99 | 131 | 141 | 22 | 40 | 44 |
| Общее число семейств | 38 | 44 | 45 | 18 | 28 | 28 |
| Среднее число видов в роде | 1,9 | 2,5 | 2,6 | 1,7 | 2,1 | 2,1 |
| Среднее число видов в семействе | 5,0 | 7,5 | 8,1 | 2,1 | 3,0 | 3,3 |
| Среднее число родов в семействе | 2,6 | 3,0 | 3,1 | 1,2 | 1,4 | 1,6 |
| Число одновидовых родов | 67 | 71 | 76 | 15 | 23 | 25 |
| Доля одновидовых родов, % | 67,7 | 54,2 | 53,9 | 68,2 | 57,5 | 56,8 |
| Число одновидовых семейств | 12 | 9 | 10 | 10 | 11 | 12 |
| Доля одновидовых семейств, % | 31,6 | 20,5 | 22,2 | 55,6 | 39,3 | 42,9 |
| Число однородовых семейств | 19 | 19 | 20 | 15 | 20 | 21 |
| Доля однородовых семейств, % | 50,0 | 43,2 | 44,4 | 83,3 | 71,4 | 75 |
| Доля видов в 5 вед. семействах, % | 47,6 | 38,3 | 38,9 | 54,1 | 65,1 | 57,1 |
| Доля видов в 10 вед. семействах, % | 67,0 | 62,1 | 61,9 | 73,0 | 74,7 | 79,1 |
| Доля видов в 20 вед. семействах, % | 86,9 | 84,3 | 83,6 | 100 | 92,8 | 96,7 |
Примечание. Районы исследования: 1 – Башкирское Предуралье, 2 – Южный Урал.
Лидерство семейства Grimmiaceae подчеркивает горный характер изученной бриофлоры и широкое распространение в регионе карбонатосодержащих пород. Высокий удельный вес семейства Pottiaceae, считающегося аридным, отмечен и во флоре мохообразных Сибири, где его присутствие объясняется широким развитием степных и лесостепных ландшафтов [Бардунов, 1992]. Очевидно, эта особенность характерна и для бриофлоры РБ. Бореальный характер флоры определяет наличие в головном спектре семейств Brachytheciaceae, Sphagnaceae, Amblystegiaceae, Dicranaceae, Bryaceae и родов Sphagnum, Dicranum, Bryum.
Ведущие семейства флоры печеночников: Scapaniaceae s.l. (29 видов), Ricciaceae (7), Lophocoleaceae (6), Cephaloziellaceae (5), Cephaloziaceae (5), ведущие роды: Riccia (7 видов), Scapania (7), Chiloscyphus (6), Cephaloziella (5), Lophozia (5), Cephalozia (5), Calypogeia (4). Высокая пропорция видов семейства Scapaniaceae является характерной чертой северных гепатикофлор Арктики и Гипоарктики [Константинова, 1989]. В то же время, представители таких семейств, как Ricciaceae, Lejeuneaceae, Metzgeriaceae, Porellaceae подчеркивают наличие в составе изученной гепатикофлоры неморальных видов.
По сравнению с горно-лесными районами Южного Урала, бриофлора Башкирского Предуралья намного беднее: на 39 % в отношении листостебельных мхов и на 50 % – в отношении печеночников (таблица 4). Видовое богатство мохообразных возрастает в горных районах и снижается на континентальных равнинах [Ignatov, 2001; Игнатов и др., 2009]. Данные по бриофлоре РБ подтверждают эту закономерность. Кроме того, отрицательное воздействие на разнообразие бриофлоры Башкирского Предуралья оказывает высокая степень хозяйственной освоенности этой территории.
В целом, таксономический анализ изученной бриофлоры показал, что по составу спектров ведущих семейств бриофлора РБ сходна с типичными бриофлорами бореальных областей, но имеет свою специфику, отражающую ее горный характер, распространение карбонатосодержащих пород и лесостепных ландшафтов. По спектру маловидовых семейств, позволяющих в определенной мере судить об истории флоры и растительности региона, изученная бриофлора имеет выраженные неморальные черты. Также следует отметить, что распространение видов, считающихся третичными реликтами, связано, главным образом, с районами Южного Урала и Уфимского плато.
Географический анализ
Отнесение видов мохообразных к тому или иному флорогенетическому зональному элементу вызывает значительные затруднения [Игнатов, 1996], поэтому в работе под географическими элементами флоры понимаются элементы только ареалогического (географического), а не зонально-генетического анализа.
Рассматривались географические элементы флоры, ранее предложенные в работах А.С Лазаренко [1956], Н.А. Константиновой [2000], М.С. Игнатова [1993], M.Hill & C.D. Preston [1998]: плюризональный, арктомонтанный, арктобореальномонтанный, бореальный, бореально-неморальный, неморальный, монтанный, аридный. Кроме того, была выделена группа видов с неясным распространением. Учитывались следующие типы ареалов: плюрирегиональный, голарктический, омниголарктический, европейский, евразиатский, евро-американский, амфиокеанический, атлантический. Отдельно была выделена группа видов с дизъюнкцией другого типа, в определении типа ареала которых возникли затруднения. Результаты анализа представлены на таблице 6.
Анализ географических элементов бриофлоры лесной и лесостепной зон РБ показал, что виды арктобореальномонтанного элемента составляет 37 %, бореального элемента – 9 %, бореально-неморального элемента – 10 %, неморального элемента – 6 %. 24 % видов являются плюризональными. Арктомонтанный, монтанный и аридный элементы представлены незначительно – по 4-5 %.
Анализ долготного распространения видов выявил высокую долю участия голарктических (45 %), омниголарктических (19 %) и плюрирегиональных (26 %) видов. Евразиатский типа ареала имеет 21 вид (5 % от всей изученной бриофлоры), европейский – 6 видов, европейско-американский 7 видов. По 2 вида имеют преимущественно амфиокеанический (Harpanthus flotovianus, Odonthoschisma denudatum) и атлантический (Cepahozia loitlesbergeri и Cephaloziella elachista) типы ареала, и, безусловно, на территории республики являются реликтовыми.
Таблица 6 Распределение видов бриофлоры лесной и лесостепной зоны РБ по географическим элементам и типам ареалов
| Геоэлементы | Типы ареалов | Всего | ||||||||
| П | Г | О | Е | Еа | Е-А | Амф | Атл | Н | ||
| Плюризональный | 73 | 12 | 20 | 4 | - | 1 | - | - | - | 110 |
| Арктомонтанный | 2 | 16 | 3 | - | - | 1 | - | - | 1 | 23 |
| Арктобореально-монтанный | 30 | 94 | 35 | - | 6 | 2 | 1 | - | 2 | 170 |
| Бореальный | 2 | 31 | 3 | 1 | - | - | - | 2 | - | 39 |
|
Бореально- неморальный |
3 | 27 | 5 | - | 8 | 2 | - | - | - | 45 |
| Неморальный | 2 | 12 | 7 | 1 | 3 | 1 | 1 | - | 1 | 28 |
| Монтанный | 3 | 8 | 6 | - | 2 | - | - | - | 1 | 20 |
| Аридный | 4 | 4 | 7 | - | 1 | - | - | - | 1 | 17 |
| Виды с неясным распространением | - | - | - | - | - | - | - | - | 4 | 4 |
| Всего: | 119 | 204 | 86 | 6 | 20 | 7 | 2 | 2 | 10 | 456 |
Примечание. Типы ареалов: П – плюрирегиональный, Г – голарктический, О – омниголарктический, Е – европейский, Еа – евразиатский, Е-А – европейско-американский, Амф – амфиокеанический, Атл – атлантический, Д с дизъюнкцией другого типа.
Бриофлора лесной и лесостепной зон РБ имеет комплексный характер. Примерно на четверть она состоит из плюризональных видов, встречающихся в разных зонах обоих полушарий, наполовину – из холодостойких видов, распространенных преимущественно в бореальной зоне и заходящих в Арктику, доля более теплолюбивых неморальных, бореально-неморальных и аридных видов не столь значительна и составляет всего одну шестую часть от выявленной флоры. Почти половина видов имеет голарктическое распространение, одна пятая часть – широко распространены в пределах Голарктики, но рассеянно встречаются в других регионах мира, евразиатский ареал имеют только 6 % видов.
Глава 8. Редкие виды мохообразных РБ и вопросы их охраны
К редким видам, встреченным не более чем в 5 местонахождениях, можно отнести 132 вида листостебельных мхов и 43 вида печеночников, что составляет, соответственно, 36 % и 47 % всей выявленной флоры листостебельных мхов и печеночников РБ.
40 % редких видов республики встречаются на наскальных местообитаниях (в том числе 5 % - в горах выше границы леса), 16 % - характерны для болот, 18 % - для лесных сообществ, 9 % - для почвенных обнажений в остепненных сообществах, по 5 % - для стоячих и медленнотекущих водоемов, а также для горных ручьев. Единичные виды отмечены на лугах и животных останках. Более 40% видов имеют дизъюнктивные ареалы (в основном арктоальпийские виды), около 25 % - рассеянное распространение, 8 % видов находятся на границе ареала.
Среди редких видов РБ верхоплодные мхи (73 %) преобладают над бокоплодными (27 %), а однодомные (51 %) - над двудомными (44 %), ложнооднодомными (4 %) и многодомными (1 %). Видов, спороношение которых в средней части Европейской России неизвестно – 27 %, редко спороносящих – 26 %. Среди однодомных наблюдается резкое преобладание видов, часто образующих спорогоны - (88 %), а среди двудомных – видов, спороношение которых в средней части Европейской России происходит редко или отсутствует.
В соответствии с системой эколого-фитоценотических стратегий (ЭФС) бриофитов, предложенной During [1979; 1992], 34% редких видов мохообразных РБ имеют стратегию челноков (с короткой продолжительностью жизни 8%, многолетних – 26%), 33% – колонистов, 33% – многолетних стайеров. Среди редких видов РБ доля колонистов ниже, а процент многолетних стайеров выше, чем, например, в Великобритании [Longton, Hedderson, 2000] или Венгрии [Órban, 1992]. Это связано с тем, что при изучении бриофлоры РБ основное внимание уделялось бриокомпоненту климаксовой растительности (особенно лесной), в то время как кратковременно существующие сукцессионные местообитания, к которым в мире приурочено немало глобально редких видов, не были объектом детального обследования.
В связи с тем, что доля редких видов во флоре мохообразных РБ довольно высока, особое внимание необходимо уделять отбору видов, нуждающихся в охране. В 2002 был опубликован 2 том «Красной книги РБ», включающий водоросли, мохообразные (22 вида листостебельных и 2 - печеночных мхов) и грибы. В нее вошли редкие виды, подходящие под один или несколько из следующих критериев отбора: включение в Красную книгу мохообразных Европы [Red Data…, 1995]: Brachythecium geheebii, Dicranum viride, Frullania bolanderi, Hamatocaulis vernicosus, Riccia frostii; виды с дизъюнктивным ареалом: Entodon concinnus, Fabronia ciliaris, Weissia squarrosa; виды с рассеянным распространением, которые в РБ встречаются редко, несмотря на достаточное количество подходящих местообитаний: Pelekium minutulum, Herzogiella seligeri, Rhynchostegium riparioides, Timmia megapolitana; виды, для которых Южный Урал является границей ареала: Plagiomnium confertidens, Brachythecium gehebii; виды, распространение которых на Южном Урале ограничено уникальными типами растительных сообществ или местообитаний: Sphagnum lindbergii, Sphagnum platyphyllum, Paludella squarrosa; виды, ареалы которых сокращаются под влиянием хозяйственной деятельности человека: Pelekium minutulum, Orthotrichum pallens, Hamatocaulis vernicosus, Paludella squarrosa и пр. Многие виды одновременно подходили под несколько критериев.
В РБ проведена инвентаризация локальных бриофлор ряда наименее нарушенных и ценных в ботаническом отношении особо охраняемых природных территорий (ООПТ): Южно-Уральского государственного природного заповедника, заповедника “Шульган-Таш”, Башкирского госзаповедника, национального парка “Башкирия”, водоохранных зон лесов Уфимского плато, пректируемого природного парка «Инзер», долин рек Белая, Нугуш и пр.). На настоящий момент репрезентативность действующей сети ООПТ РБ составляет 55 % для всех редких видов мохообразных республики, и 58 % - для видов, включенных в Красную книгу РБ. Для сосудистых растений этот показатель гораздо выше и составляет более 70 %. Такая ситуация свойственна многим регионам, поэтому необходимо проводить специальный учет местообитаний с уникальными комплексами бриофитов еще на этапе проектирования ООПТ, иначе их эффективность для сохранения разнообразия мохообразных будет невысока.
При охране мохообразных необходимо учитывать то, что среди редких таксонов высока доля неконкурентоспособных видов, которые могут расти только на кратковременно существующих, возникающих после нарушений местообитаниях. В РБ это приблизительно 45 % редких видов, места произрастания которых, как правило, обнаружены вне пределов ООПТ. Для сохранения их популяций необходим специальный экологический менеджмент по поддержанию определенного режима нарушений, способствующего возобновлению кратковременно существующих местообитаний.
Наиболее общим подходом для сохранения мохообразных на землях с интенсивным хозяйственным и рекреационным использованием является выявление обособленных специфических местообитаний: ручьев, скальных выходов, заболоченных участков, обнажений суглинков, периодически пересыхающих водоемов и пр., для которых характерно повышенное разнообразие и концентрация редких видов мхов [Heinlen, Vitt, 2003]. Сравнительно небольшие затраты на огораживание таких участков, прокладывание мостков и дорожек вдоль ручьев, оборудование туристических стоянок и подходов для смотровых площадок позволяют сохранить местообитания бриофитов даже в зонах с интенсивной рекреационной нагрузкой. В условиях РБ это особенно актуально для долин рек Белая, Нугуш, Инзер, служащих излюбленными местами отдыха туристов.
В РБ на настоящий момент охрана местообитаний мохообразных осуществляется только в системе ООПТ, что не является достаточным условием для сохранения редких видов этой группы растений. Требуются специальные исследования для разработки вопросов поддержания разнообразия мохообразных в зонах интенсивной рекреации и лесах хозяйственного использования.
ВЫВОДЫ
1. В сообществах 42 ассоциаций лесной растительности РБ выявлено 180 видов листостебельных мхов и 32 – печеночников. Наибольшее разнообразие мохообразных отмечено в темнохвойных и темнохвойно-широколиственных лесах союзов Piceion excelsae и Aconito-Piceion, наименьшее – в дубовых и березово-сосновых лесах союзов Lathyro-Quercion и Veronico-Pinion. При повышении роли темнохвойных пород в древостое возрастают видовое богатство мохообразных и вклад верхоплодных мхов и печеночников. Уровень флористического сходства бриоценофлор лесов разных типов довольно низкий, что объясняется преобладанием видов с рассеянным распространением и низким постоянством в сообществах. Основными факторами, определяющими распространение мохообразных, являются наличие подходящих субстратов и местообитаний, представленных в широком спектре лесных сообществ. Характерной чертой южно-уральских лесов является высокая доля эпилитных видов (от 11 до 25 % в бриоценофлорах разных союзов).
2. Во всех типах лесной растительности РБ ведущая роль принадлежит двудомным видам мохообразных (53-65 % от бриоценофлоры союзов). Виды разных субстратных групп различаются по спектрам половых типов, жизненных стратегий и активности размножения. В эпифитно-эпиксильном комплексе преобладают однодомные виды со стратегиями колонистов и челноков, характеризующихся высокой частотой спороношения. Среди эпилитных и эпигейных мохообразных доминируют двудомные виды со стратегией многолетних стайеров, редко образующих спорогоны и размножающихся в основном вегетативно. Полное выявление бриоценофлор и учет спектра жизненных стратегий видов может быть использовано для оценки стабильности экологического режима лесного сообщества и длительности их ненарушенного существования.
3. Выделены группы видов мохообразных-индикаторов биологически ценных лесов региона. Для индикации старовозрастных лесов чаще всего используют эпифиты и эпиксилы, требовательные к качеству субстрата, но имеющие довольно высокую активность размножения. Индикаторами девственных лесов могут служить только эпигейные и эпилитные реликтовые виды с низкой активностью размножения. На территории РБ эти виды чаще всего встречаются в глубоко врезанных долинах горных рек Уфимского плато и центрально-возвышенной части Южного Урала.
4. Бриосинтаксономия РБ включает сообщества 16 ассоциаций, 9 союзов, 6 порядков и 4 классов. По сравнению с синтаксонами, выделенными в Западной и Центральной Европе, эпифитные и эпиксильные бриосообщества РБ имеют сильно обедненный флористический состав, что вызвано континентальностью климата Южного Урала. Прибрежно-водные брисообщества обладают высокой степенью сходства с европейскими аналогами.
5. Флора мохообразных лесной, лесостепной зон РБ и их горных аналогов насчитывает 365 видов листостебельных мхов и 91 вид печеночников. По сравнению с равнинными районами Башкирского Предуралья, разнообразие мохообразных Южного Урала выше на 39 % в отношении листостебельных мхов, на 50 % – печеночников. По таксономическому составу выявленная флора сходна с типичными бриофлорами бореальных областей, но имеет свою специфику, отражающую горный характер, распространение карбонатосодержащих пород и наличие лесостепных ландшафтов. Более половины видов имеет голарктическое распространение, одна пятая часть – широко распространены в пределах Голарктики, но рассеянно встречаются в других регионах мира, одну четвертую часть составляют плюризональные виды, распространенные в разных зонах обоих полушарий, евразиатский ареал имеют 6 % видов.
6. Во флоре РБ 132 вида листостебельных мхов (36 %) и 43 вида печеночников (47 %) являются редкими. Среди них значительную часть представляют виды с дизъюнктивными ареалами и рассеянным распространением. Охрана мохообразных РБ осуществляется только в системе ООПТ, что не является достаточным условием для сохранения редких видов этой группы. Требуются специальные мероприятия по охране разнообразия мохообразных в зонах интенсивной рекреации и лесах хозяйственного использования.
Список основных работ, опубликованных по теме диссертации
Статьи в научных журналах, рекомендованных ВАК МОН РФ для защиты докторских диссертаций
1. Баишева Э.З., Соломещ А.И. Бриосинтаксономия: эпифитные и эпиксильные сообщества // Бюллетень МОИП. Отд. биол. 1994. Т. 99, вып. 6. С. 74-85.
2. Баишева Э.З., Потемкин А.Д. К флоре печеночных мхов Башкирии // Ботанический журнал. 1998. Т. 83, № 9. C. 46-51.
3. Мартыненко В.Б., Жигунов О.Ю., Баишева Э.З., Журавлева С.Е., Миркин Б.М. Экологическое разнообразие заповедника «Шульган-Таш» // Бюллетень МОИП. Отд. биол. 2003. Т.108, вып. 5. С. 32-39.
4. Мартьянова С.Н., Мартыненко В.Б., Жигунова С.Н., Баишева Э.З., Журавлева С.Е., Миркин Б.М. Экологическое и биологическое разнообразие водоохранных лесов Павловского водохранилища // Бюллетень МОИП. Отд. биол. 2004. Т.109, вып.4. С. 50-57.
5. Миркин Б.М., Мартыненко В.Б., Баишева Э.З., Журавлева С.Е., Мулдашев А.А., Широких П.С. Вклад сохранения биологического разнообразия в экологическую безопасность // Безопасность жизнедеятельности. 2006. № 10. С. 36-39.
6. Баишева Э.З. Разнообразие мохообразных естественных экосистем: подходы к изучению и особенности охраны // Успехи современной биологии. 2007. Т.127, № 3. С.316-333.
7. Баишева Э.З., Жигунова С.Н., Мартыненко В.Б., Широких П.С. Экологические и фитоценотические особенности бриокомпонента водоохранных лесов Уфимского плато // Экология. 2009. № 3. С. 1-7.
8. Баишева Э.З. Игнатова Е.А. Новые виды мхов в Республике Башкортостан // Ботанический журнал. 2009. Т.94, № 5. С.138-140.
9. Баишева Э.З. К бриофлоре пойменных сероольшаников Республики Башкортостан // Вестник ОГУ. 2009. № 6 (100). С. 50-53.
10. Chytrỳ M., Danihelka J., Horsák M., Kočí M., Kubešová S., Lososevá Z., Otỳpková Z., Tichỳ L., Martynenko V.B., Baisheva E.Z. Modern analogues from the Southern Urals provide insights into biodiversity change in the early Holocene forests of Central Europe // Journal of Biogeography. 2010. V. 37. P. 767–780.
Статьи в центральных российских изданиях
1. Baisheva, E.Z., Solometch, A.I., Ignatova E.A. Bryophyte vegetation of Bashkiria, South Urals. I. Epiphytic and epixylic communities // Arctoa. 1994. V. 3. P. 139-159.
2. Baisheva, E.Z. Bryophyte vegetation of Bashkiria, South Urals. II. Сommunities of north-eastern Bashkiria // Arctoa. 1995. V. 4. P. 55-63.
3. Baisheva, E.Z. Bryophyte vegetation of Bashkiria, South Urals. III. Epiphytic and epixylic communities of western Bashkiria // Arctoa. 2000. V. 9. P. 101-104.
4. Баишева Э.З. Дополнение к бриофлоре Башкирии (Южный Урал) // Новости систематики низших растений. 2002. Т.36. С. 210-212
5. Золотов В.И., Баишева Э.З. Флора листостебельных мхов заповедника «Шульган-Таш» (Республика Башкортостан, Россия) // Arctoa. 2003. V. 12. C. 121-132.
6. Баишева Э.З., Соломещ А.И., Григорьев И.Н. Обзор растительности ручьев Южного Урала // Растительность России. 2004. № 6. C. 3-14.
7. Журавлева С.Е., Соломещ А.И., Баишева Э.З. Сообщества эпифитных лишайников с Lobaria pulmonaria (L.) Hoffm. в старовозрастных лесах на Южном Урале // Растительность России. 2004. № 6. C. 15-22.
8. Красногорская Н.Н., Баишева Э.З., Вдовина И.В. Мониторинг содержания тяжелых металлов в атмосфере г.Уфы с использованием листостебельных мхов // Безопасность жизнедеятельности. 2007. № 9. С. 32-36.
9. Баишева Э. З., Мулдашев А. А., Игнатова Е. А. К флоре мохообразных проектируемого природного парка «Инзер» (Южный Урал) // Новости систематики низших растений. 2009. Т. 43. С. 297-312.
10. Потемкин А.Д., Баишева Э.З. Новые находки печеночников в Республике Башкортостан. 1 // Arctoa. 2009. V.18. P. 259-260.
Монографии
1. Красная книга Республики Башкортостан (объединенный том) / Под ред. А.А.Фаухутдинова. Уфа: Полипак, 2007. 528 с.
2. Водоохранно-защитные леса Уфимского плато: экология, синтаксономия и природоохранная значимость / Кол. авторов. Под ред. А.Ю. Кулагина. Уфа: Гилем, 2007. 448 c.
3. Флора и растительность Южно-Уральского государственного природного заповедника / Кол. авторов. Под ред. Б.М. Миркина. Уфа: Гилем, 2008. 528 с.
Статьи в крупных региональных изданиях и сборниках трудов
1. Баишева, Э.З., Соломещ, А.И., Игнатова, Е.А. Синтаксономия эпифитной и эпиксильной моховой растительности Башкирии. Уфа: Институт биологии БНЦ УрО РАН, 1993. 49 с. (Рукопись деп. В ВИНИТИ 18.11.93, №2854-В93).
2. Соломещ, А.И., Григорьев, И.Н., Хазиахметов, Р.М., Баишева, Э.З. Синтаксономия лесов Южного Урала.V. Хвойно-широколиственные леса. Уфа: Институт биологии БНЦ УрО РАН, 1993. 68 с. (Рукопись деп. В ВИНИТИ 02.06.93, №1464-В93).
3. Баишева Э.З., Игнатова, Е.А. Материалы к флоре эпифитных и эпиксильных мхов Республики Башкортостан // Вопросы рационального использования и охраны растений в Республике Башкортостан: сборник статей. Уфа: Гилем, 1998. С.156-164.
4. Гапон С.В., Баïшева Е.З. Еколого-флористична характеристика брiоугруповань вiльхових ценозiв Полтавщини та особливостi ïх класифiкацiï // Збiрник наукових праць Полтавського державного педагогiчного унiверситету iменi В.Г. Короленко. Випуск 3 (24), серiя «Екологiя. Бiологiчнi науки». 2002. С.30-34.
5. Соломещ А.И., Мартыненко В.Б., Баишева Э.З., Журавлева С.Е., Онищенко Л.И. Характеристика флоры и растительности в зоне влияния Юмагузинского водохранилища и пути ее сохранения // Экологические аспекты Юмагузинского водохранилища. Уфа: Гилем, 2002. С. 93-113.
6. Ямалов С.М., Мартыненко В.Б., Голуб В.Б., Баишева Э.З. Продромус растительных сообществ Республики Башкортостан: Препринт. Уфа: Гилем, 2004. 64 с.
7. Баишева Э.З. О редких видах листостебельных мхов Республики Башкортостан // Материалы Международного совещания «Актуальные проблемы бриологии», посвященного 90-летию со дня рождения А.Л. Абрамовой. СПб, 2005. C.19-25.
8. Баишева Э.З. Предварительный список мохообразных Национального парка «Башкирия» // Экологические аспекты сохранения биологического разнообразия Национального парка «Башкирия» и других территорий Южного Урала: Сборник научных статей. Уфа: Информреклама, 2007. С.63-65.
9. Баишева Э.З. Мохообразные в растительном покрове Башкортостана // Вестник Академии наук РБ. 2008. Т.13, № 2. С.31-37.
Патент Российской Федерации
Патент на изобретение № 2321030 «Способ оценки содержания тяжелых металлов в атмосферном воздухе с помощью листостебельных мхов» (2008 г.). Авторы: Красногорская Н.Н., Вдовина И.В., Баишева Э.З., Минуллина Г.Р.
© 2010 Интернет База Рефератов