
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Курсовая работа: Стійкість до голодування і активність АДГ у Drosophila melanogaster із природних популяцій України
Курсовая работа: Стійкість до голодування і активність АДГ у Drosophila melanogaster із природних популяцій України
Курсова робота
Стійкість до голодування і активність АДГ у Drosophila melanogaster із природних популяцій України
Реферат
Дослідження проводились в 2008 – 2009 роках на кафедрі генетики та молекулярної біолог ОНУ ім. І.І. Мечникова.
Метою досліджень було вивчення тривалості життя в умовах голодування та визначення активност алкогольдегідрогенази у мух із природних популяцій України.
Встановлено, що самки із популяцій Одеси, Варви і Пирятина були більш витривалими в умовах голодування порівняно з самцями. Найбільшу стійкість до голодування виявили мухи Одеської популяції, а найменшу мухи Київської популяції. стійким до голодування мухам із Одеської популяції притаманна досить висока активність АДГ, а чутливим до голодування мухам Київської популяц менша активність ферменту.
В цілому в умовах проведеного експерименту не виявлено прямо залежності між витривалістю мух в умовах голодування і активністю їх АДГ.
Зміст
Вступ
1. Огляд літератури
1.1 Стійкість до голодування як показник пристосованості
Drosophila melanogaster
1.2 Активність алкогольдегідрогенази у Drosophila melanogaster
2. Матеріали та методи досліджень
3. Результати досліджень та х обговорення
Узагальнення
Висновки
Список цитовано літератури
Вступ
Концепція пристосованості організмів до оточуючого середовища є однією із центральних в генетиці популяцій та в теор еволюції.
Єдиною силою, що спрямовує еволюцію органічного світу, є природній добір. Ця спрямовуюча дія добору проявляється і на початкових етапах еволюційних змін, що відбуваються в популяціях. Поняття ”пристосованість”, яке використовував ще Ч. Дарвін в якост кількісної міри природного добору або міри ефективності розмноження генотипу, зараз набуло більш широкого змісту. Ймовірність дати нащадків визначається багатьма властивостями організму – його життєздатністю, швидкістю досягнення репродуктивного віку, здатністю до схрещування, стійкістю до голодування, плодючістю. Сукупність цих властивостей називається пристосованістю особини до умов середовища, в якому вона існує [Айала, Кайгер, 1987].
Пристосованість часто ототожнюють з адаптацією, при цьому пристосованість являється більш універсальним поняттям, що включає в себе адаптацію як складовий компонент. Під адаптацією прийнято розуміти складний комплекс захисних і пристосувальних реакцій (специфічних і неспецифічних), що обумовлені різними генетично детермінованими механізмами (фізіологічними, біохімічними, морфологічними).
Адаптація може бути модифікаційною, яка дозволяє організму в межах норми реакції залишатися життєздатним і давати потомство в нових умовах, та генотипічною, при якій перебудовується структура генотипу і утворюються нов норми реакції. Генотипічна адаптація відбувається в процесі природного та штучного добору і забезпечує пристосування організмів до нових умов існування. Взаємодія вказаних двох систем адаптації забезпечує максимальну пристосованість живих організмів до мінливих умов навколишнього середовища.
Однією із показових характеристик пристосованості є стійкість до голодування здатність вижити в екстремальних умовах нестачі корму. Так як нші фенотипічні характеристики пристосованості, стійкість до голодування в значній мірі визначається генотипом, контролюється великою кількістю взаємодіючих генів і проявляє значну мінливість в природних популяціях. Ось чому генотипічно різним особинам зазвичай притаманна неоднакова стійкість до голодування.
Улюбленим об’єктом генетичних досліджень є плодова мушка Drosophila melanogaster – найбільш вивчений в цьому плані вид дрозофіли. Результати тривалого дослідження дрозофіли дають можливість проаналізувати різноманіття всього спектру мутацій, характер розподілу генів по різних хромосомах, співвідношення домінантних та рецесивних мутацій та ін. Ці дані являють собою загальногенетичну цінність дозволяють прогнозувати генетичні ситуації, з якими ймовірно можна зустрітися у інших вищих організмів. Використання в якості об’єкта генетичних досліджень плодової мушки дрозофіли привело до значних успіхів в цій галузі [Тоцький, Хаустова та ін., 2002].
Більшість тварин різних видів на протязі життя переживають періоди скорочення раціону і тому у них еволюційно закріплені адаптації, що підсилюють х стійкість до голодування. Знання про генетичні та фізіологічні засади цих адаптацій, їх еволюційні зв`язки (як внутрішньовидові, так і популяційні) отримані значною мірою завдяки дослідженням на дрозофілі. Так відомо, що окрем мухи можуть направлено посилювати стійкість до голодування у відповідь на легкий харчовий стрес (Harshmann,1998).
Зважаючи на вищезазначене, метою роботи було вивчення тривалості життя в умовах голодування та визначення активност алкогольдегідрогенази у мух із природних популяцій України.
Згідно із метою були визначені наступні завдання:
1). В умовах голодування дослідити тривалість життя та визначити статеві особливості стійкості до голодування у мух досліджуваних популяцій.
2). Визначити активність АДГ у мух досліджуваних популяцій.
Об’єкт дослідження − пристосованість Drosophila melanogaster.
Предмет дослідження − стійкість до голодування та активність АДГ у Drosophila melanogaster різного походження.
1. Огляд літератури
1.1 Стійкість до голодування як показник пристосованості Drosophila melanogaster
Тривалість життя дорослої мухи з моменту її вилуплення із лялечки в лабораторних умовах складає 3 – 4 тижні та в значній мірі залежить від умов утримання (температури, вологості, якості корму, густоти заселення, наявност бактерій). В спеціальних дослідах дрозофіла жила до 153 днів (Медведєв, 1966). Самки, як правило, мають більшу тривалість життя, ніж самці, що може бути викликане генетичними та фізіологічними причинами.
Справді, про більшу тривалість життя самок було сказано давно. Ще Дарвін розглядав меншу тривалість життя самців як “природню конституційну властивість, що обумовлена статтю” (Гаврилов, 1991). Таким чином факти про більшу життєздатність особин жіночої статі мають право на існування.
Відмінності в тривалості життя в межах однієї статі, як правило, можуть бути зумовлені фізіологічними причинами. Так, віргінні самки, мають більшу тривалість життя, ніж запліднені (Зіміна, Малиновський, 1977). Згідно із літературними даними, направленість статевих відмінностей в резистентності до голодування є генетично детермінованою (Хаустова, 1995). Вивчення тривалост життя самок та самців дрозофіли в умовах харчового стресу – цікава і водночас важлива проблема сучасної генетики та науки в цілому.
Стресом можна назвати певний фактор середовища, внаслідок д якого послаблюється стійкість організму до даного чинника. Якщо організм зміг вижити після дії стресового фактора, то у нього виробляються специфічні адаптації, що пом’якшують наслідки при повторній його дії (Haffman, Parsons, 1991; Randall, 1997). Однією із найбільш поширених причин харчового стресу у тварин, у тому числі і у дрозофіли, є скорочення кількості їжі або її недостатня якість.
Відомо, що енергетичні процеси відіграють центральну роль в організмі, тому голодування провокує зміни на різних рівнях прояву фенотипу – від змін внутрішньоклітинних сигналів – до зсувів фаз життя в онтогенезі.
Існує думка, що збільшена тривалість життя потребує від організмів певних змін, які, скоріш за все, взаємопов`язані і з іншими характерними рисами пристосованості виду. Отже, природний добір повинен підтримувати ті генотипи, що здатні змінювати свої фізіологічні особливості для підвищення стійкості до голодування у відповідь на сигнали, що сповіщають про початок періоду голодування. Іншими словами, очікується, що дана ознака в деякій мірі покаже рівень адаптивної фенотипічної пластичності (Partridge, 2005).
Встановлено, що скорочення кількості їжі (в основному дріжджів) для дорослих мух, призводить до збільшення їх витривалості майже в два рази в порівнянні з тими, які взагалі не отримували дріжджі з їжею (Chippindale et al., 1998; Leroi et al., 1994; Kapahi et al., 2004; Piper et al., 2005; Burger et al., 2007). В той же час показано, що у мух, які піддавались надто тривалому обмеженню раціону, виживаність навпаки знижувалась (Burger et al., 2007).
Відомо, що підвищена стійкість до голодування та до інших чинників стресу визначає стан особини під час діапаузи, яка може бути викликана низькими температурами або коротким фотоперіодом. Це означає, що певній особині буде легше витримати період діапаузи, якщо у неї виробилась висока витривалість під час голодування, пережитого раніше.
Варто також зазначити, що механізми фізіологічних реакцій на харчовий стрес зберігаються і закріплюються у особин усього тваринного світу (Tatar; 2003; Partridge et al., 2005; Arsham, Neufeld, 2006). Таким чином, дані щодо розуміння генетики і фізіології адаптації до голодування, отримані в експериментах на дрозофілі, можна застосовувати не лише до мух, а й до комах та деяких інших тварин взагалі.
Практично у всіх дослідженнях, що стосуються вивчення стійкості до голодування Drosophila melanogaster, мова йде про гостре голодування, тобто повне позбавлення їжі (Huey, 2004). Але в роботах деяких авторів показано, що мухи дрозофіли також дуже чутливі до нестачі води, адже існу досить тісний зв`язок між стійкістю до голодування та здатністю витримати нестачу вологи (Partridge, Piper, 2005). Виходячи із наведених даних автори рекомендують вносити воду у вигляді вологих пробок чи смужок фільтрувального паперу до середовища, де голодують мухи. Отже, цю рекомендацію варто враховувати при постановці експериментів.
Зазвичай тривалість життя особин за голодування визначають як час, що проходить до загибелі 50% (Lt 50) мух в умовах досліду. В залежності від генотипу та умов середовища, середній час виживання без корму коливається від 20 годин для самців і до більш ніж 50 годин для самок (Harshman , Shmidt, 1998; Harshmann, 1999; Baldal, 2005; Harbison, 2005).
Еволюційний взаємозв’язок між голодуванням та тривалістю життя був підтверджений деякими експериментами з використанням методів селекції. Дан відносно дрозофіл отримані головним чином в дослідженнях на Drosophila melanogaster. Досліджувані лабораторні лінії дрозофіли, відібрані за ознакою тривалості життя, за даними деяких авторів (Rose, Archer, 1996) демонстрували відносно кращу стійкість до голодування. За даними інших авторів (Rose et al., 1992) в свою чергу, селекція на стійкість до голодування в умовах експерименту призводила до збільшення тривалості життя. В ході досліджень також було з’ясовано, що мутанти Drosophila melanogaster з великою тривалістю життя зазвичай є стійкими і до інших чинників стресу (Lin, Benzer, 1998).
Добре відомо, що гени всіх живих організмів кодують інформацію для синтезу білкових молекул, із яких створюються всі компоненти клітин живих організмів та які приймають участь у каталізі всіх біохімічних процесів. Фактично кожну біохімічну функцію чи елемент тіла у дрозофіли вдалося пов’язати з дією того чи іншого гена (Тоцький, 2002). Усестороннє розуміння еволюційних механізмів становлення стійкості до голодування потребує ідентифікації та характеристики генних локусів, що вносять вклад в спадкову мінливість ц ознаки та лежать в основі її еволюційних змін. Мутації або ж маніпуляції з експресією генів, що посилюють витривалість до голоду, вказують на можливост змін в штучних умовах, але досі залишається незрозумілим чи мають встановлені зміни місце у природі.
Певне розуміння генетичних аспектів, а саме успадковування та мінливість стійкості до голоду, може бути отримане шляхом аналізу схрещувань між стійкими та нестійкими генотипами. Вчені використали цей підхід для вивчення генетичної структури відмінностей між двома парами географічно віддалених популяцій дрозофіли із Південної Америки та Австралії. Цікавим виявилось те, що обидві статі мух у Південній Америці та самки дрозофіл у Австралії виявляли позитивні ефекти домінування по материнській лінії, тобто нащадки виявляли вищу стійкість до голодування, якщо ця ознака була високою у матері.
Разом із простим материнським ефектом були виявлені також парадоксальні результати, а саме: менш стійкі до голоду матері давали більш стійких нащадків обох статей. Причини виявлених закономірностей і дос залишаються невідомими.
Результати, отримані при картуванні генів кількісних ознак, а також дані тестування на кількісну комплементацію (Mackay, Fry, 1996) дозволили визначити шлях до ідентифікації специфічних локусів, що відповідають за неоднакову стійкість до голодування. Як випливало із попередніх даних, природний поліморфізм локусу десатурази-2 міг впливати на стійкість мух до голодування (Greenberg et al., 2003), але подальші експерименти (Coyne, Elwyn, 2006) не підтвердили це припущення. Використовуючи більш комплексний підхід, було ідентифіковано 13 локусів (6 із них мали статево-специфічний ефект), які вносять вклад в розбіжності щодо стійкості до голодування між двома лабораторними лініями мух. Ці локуси включають гени, що беруть участь в оогенезі (ген l(2)G270 впливає на розвиток яйцеклітини) та метаболізмі (гени, що регулюють розподіл жирів). Також були визначені гени, що впливають на харчову поведінку мух (наприклад, ген NaСР60Е).
Вченими також вивчалися зміни в експресії генів, викликані умовами голодування. Даний підхід не є досить інформативним щодо генетичної мінливост здатності витримати голод, але проливає світло на молекулярні механізми відповіді на харчовий стрес. При голодуванні гени, що задіяні в біосинтез білків і гідролазній активності, мають тенденцію до неврегульованості та надлишкового синтезу. Таким чином організм намагається компенсувати нестачу поживних речовин і, як наслідок, пережити несприятливі умови голодування.
Відмінності в ході добору по стійкості до голодування можуть призводити до відмінностей за цією ознакою між популяціями. Докази таких відмінностей знайдені при вивченні великих географічно віддалених популяцій. Вченими було показано, що на Індійському півострові має місце негативна кореляція між стійкістю до голодування та географічною широтою для п’яти видів дрозофіли (включаючи і Drosophila melanogaster). Аналогічним чином, розподіл по довготі був недавно показаний для двох інших видів дрозофіли на Індійському півострові (Parkash, 2005). Навпаки, позитивна кореляція між досліджуваною ознакою та широтою у Drosophila melanogaster, була виявлена на сході Південної Америки (Schmidt, 2005). В той же час ніяких варіацій по даній ознаці не було виявлено у особин з Південної Америки та Східої Австралії. Таким чином, наведені дані свідчать про те, що залежність між стійкістю до голодування та географічною широтою підтверджується не завжди.
Аналізуючи дані літератури щодо стійкості до голодування особин популяції Drosophila melanogaster, можна відзначити, що за останні роки були проведені дослідження, спрямовані на розкриття молекулярних та фізіологічних механізмів відповіді на харчовий стрес. Існує думка, що дана ознака відтворює рівень адаптивної пластичності та являється частиною механізму виживання, який може частково піддаватися інсуліновій системі регуляції.
Що стосується розуміння екологічних аспектів витривалості до голодування, то і досі залишається багато питань відносно природного добору за цією ознакою.
Дрозофіла надає унікальну можливість для повноцінного розуміння та нтеграції різних аспектів еволюційної відповіді на харчовий стрес.
1.2 Активність алкогольдегідрогенази у Drosophila melanogaster
Механізми адаптації генотипів та популяцій до дії екологічних факторів є досить цікавими і тому інтенсивно вивчаються в багатьох лабораторіях. В даному контексті вважається доцільним вияснити роль ферменту алкгольдегідрогенази (АДГ) в життєдіяльності та адаптації у Drosophila melanogaster. Ген-ензимна система АДГ на протязі тривалого часу притягу увагу численних дослідників в різних областях генетики – від молекулярної до популяційної, завдяки відносно простій ідентифікації ферменту, значному поліморфізму тій ключовій ролі, що АДГ відіграє в життєдіяльності дрозофіли (цей фермент допомагає здійснювати детоксикацію та утилізацію спирту, який являється важливим компонентом середовища існування плодової мушки).
Фермент АДГ (по класифікації ферментів – КФ 1. 1. 1. 1.) відноситься до класу оксидоредуктаз, об`єктом дії яких є група СН-ОН.
Ацетальдегід + НАДН + Н+ → етанол + НАД
Алкогольдегідрогеназа широко розповсюджена в природі. Алкогольдегідрогеназна активність притаманна різним клітинам всіх живих організмів.
Піридиновий нуклеотид в якості коферменту відіграє головну роль в реакції з усіма вивченими АДГ, при цьому фермент може окислювати не лише етанол, але й інші первинні та вторинні спирти [Діксон, 1982]. При перетворенн етилового спирту в ацетальдегід спостерігається впорядкована багатоточкова взаємодія в АДГ між ферментним білком, субстратом і коферментом.
АДГ дрозофіли складається із двох субодиниць з молекулярною масою 60 кДа та відрізняється від інших алкогольдегідрогеназ відсутністю Zn2+. Крім цього АДГ дрозофіли проявляє неабияку спорідненість до вторинних спиртів. Швидкість реакції, що каталізується АДГ Drosophila melanogaster, при використанні вторинних спиртів як субстратів, в декілька разів більша, ніж швидкість окислення етанолу. Амінокислотна послідовність АДГ дрозофіли відрізняється від інших алкогольдегідрогеназ. При картуванні пептидів АДГ Drosophila melanogaster виявлена наявність активного залишку цис-135 в домені, що зв'язує НАД+, а також двох залишків амідів на С-кінці пептиду. Таким чином, буде відрізнятися також і механізм дії АДГ дрозофіли – даний фермент формує нестійкий комплекс із субстратом і коферментом [Chambers, 1984].
Алкогольдегідрогеназа відіграє головну роль в каталізі останнього етапу спиртового бродіння, що притаманне дріжджам, а також тканинам, що знаходяться в анаеробному стані [Ленінджер,1985; Страєр, 1985]. Для АДГ плодової мушки більш характерною є зворотна реакція окислення спиртів до альдегідів чи кетонів, оскільки спирти є важливим компонентом середовищ снування дрозофіли. Drosophila melanogaster відрізняється від інших видів дрозофіли здатністю використовувати етанол та інші спирти як джерела поживних речовин на різних стадіях свого розвитку. АДГ є основним ферментом в метаболізмі етилового спирту у мух. Мутанти, яким не притаманна алкогольдегідрогеназна активність, є дуже чутливими до токсичної дії спирту та не можуть використовувати його [Economos, 1986 ; McKechnie , 1984].
Більш детальне вивчення системи метаболізму етанолу у D. melanogaster та деяких інших видів дозволило виявити наявність двох самостійних ферментів: алкогольдегідрогенази та альдегіддегідрогенази, що приймають безпосередню участь у деградації етилового спирту, а також встановити пряму кореляцію між їх активностями у дорослих особин. Це дало вагому підставу припустити, що у дрозофіли окислення етанолу здійснюється через ацетальдегід шляхом послідовної дії двох вищезазначених [Garcin, 1986, Lietart,1985]. Наявність альдегіддегідрогенази виявляється переважно у фракції мітохондрій, а алкогольдегідрогенази – в цитозолі [Lietart, 1985; Garcin, 1986].
Результати численних досліджень показали наявність в усіх природних популяціях Drosophila melanogaster двох варіантів АДГ, що відрізняються электрофоретично – АДГ-F и АДГ-S. Вони знаходяться під контролем структурного гена Adh, що локалізований в хромосомі 2 в положенні 50.1. Амінокислотні послідовност білків класу АДГ-F відрізняються від таких класу АДГ-S заміною лізину на треонін в позиції 192. Ця амінокислотна заміна торкається тієї області, яка являється каталітичним доменом ферменту, що і лежить в основі різниці у відносній активності, термостабільності та інших властивостях алозимів [Chambers, Wilks et al., 1984]. Завдяки генетичному поліморфізму, локус Adh відіграє важливу роль в генотипічній адаптації дрозофіли до факторів зовнішнього середовища.
При електрофорезі в поліакриламідному гелі АДГ виявляється в виді двох – трьох зон з головною катодною смугою (АДГ-5) та додатковими більш рухливими смугами (АДГ-3 і АДГ-1). У АДГ-S, яка є менш рухливою, смуги на електрофореграмах зміщуються ближче до катоду і займають положення 7, 5, 3 відповідно [Крутовський, 1983; Chambers, 1984; 1991; Johnson, 1964; Schwartz, 1976].
Встановлено, що при тривалому голодуванні мух, або при їх обробці низькими концентраціями ацетилацетону, проявляється високий рівень експресії АДГ-1 та АДГ-3 за рахунок конверсії алозимного спектру. Дія на мух різних концентрацій етанолу призводила до ретроконверсії молекулярних форм АДГ [Heinstra, 1986].
Припускається [Mckechnie, 1984], що взаємне перетворення модифікованих молекул АДГ, що характеризуються різною стабільністю та кінетичними характеристиками, є важливою ланкою в складній системі адаптації Drosophila melanogaster до екзогенного етанолу. Відповідно до цієї гіпотези екзогенний етанол стимулює ріст активност АДГ, збільшуючи співвідношення НАДН / НАД, тим самим знижуючи відносну кількість АДГ-1 на користь АДГ-3 та АДГ-5.
Здійснюючи регуляцію рівнів НАД / НАДН і НАДР / НАДРН, які в свою чергу регулюють розпад нших ферментів, АДГ також впливає на життєздатність дрозофіли [Xinmi, 1992]. Очевидно, такий спосіб регуляції активності фермента − найбільш важливий в процесах адаптації [Anderson,1983].
Численн дослідження показали, що здатність мух до існування в умовах з високим вмістом алкоголю знаходяться в залежності від рівня активності їх АДГ [Pecsenye, 1994; 1997]. Хоча вирішальне значення для виживання мух при збільшених концентраціях спирту відіграє абсолютна кількість АДГ, а не активність ферменту [Anderson, 1983]. Це підтверджено імунохімічними та молекулярно-біологічними дослідженнями. Так, встановлено, що кількість білку АДГ визначається швидкістю синтезу, що позитивно корелює із рівнем цитоплазматичної мРНК [Anderson,1983; Birley, 1984]. Таким чином, регуляцію активності АДГ за рахунок зміни кількості ферментного білку на різних стадіях розвитку Drosophila melanogaster можна вважати достовірно встановленою.
В зв’язку із важливою роллю ген-ензимної системи АДГ в процесах метаболізму і адаптації дрозофіли, рівень активності та інші характеристики алкогольдегідрогенази можна розглядати як критерій пристосованості генотипу.
2. Матеріали та методи досліджень
Дослідження проводили на мухах Drosophila melanogaster, що відносяться до родини Drosophilidae із відділу Diptera. Дрозофіла класичним об’єктом генетичних досліджень, завдяки наступним властивостям. По-перше, у неї порівняно короткий період онтогенезу; по-друге, вона легко виживає та розвивається в лабораторних умовах, а також не потребує значних витрат на розведення та харчування.
Для цих мух комфортною температурою вважається 24 – 25С. При такій температурі цикл розвитку дрозофіли від яйця до дорослої мухи склада приблизно 10 діб. Розвиток яйця триває 20 годин, а розвиток личинки і лялечки 8 діб. Таким чином за рік можна отримати до 40 поколінь дрозофіли. При температурі вищій за 31С дрозофіла стає безплідною, хоча в природних умовах вона може витримувати і більш високі температури. Зі зниженням температури цикл розвитку мух дуже уповільнюється. Так, при температурі 10С період личинки та лялечки значно зростає у часі.
культивування мух із досліджуваних популяцій проводили у скляних банках (200 мл) при температурі 24С у стандартних умовах. для годування дрозофіли зазвичай використовують корм із дріжджами. Крім них також використовували: цукор як субстрат для розвитку дріжджів та агар-агар, що надає корму желеподібної консистенції. Харчову масу готують із наступних співвідношень компонентів на 1 літр води:
агар-агар – 50 г
цукор-пісок – 36г
манна крупа – 36г
дріжджі – 27г
В ході експерименту аналізу піддавали мух із популяцій різного походження – К (Київ), Од (Одеса), В (Варва), П (Пирятин), Оз (Озеро із Чорнобильської зони) та сад (Яблуневий сад із Чорнобильської зони), що були люб`язно надані співробітниками кафедри генетики Київського національного університету ім. Т.Г. Шевченка.
Тривалість життя мух за голодування досліджували наступним чином. У пробірки (10 мл) без корму поміщали окремо по 10 самок та самців кожно популяції. Використовували особин триденного віку, попередньо на протязі трьох діб вони мали можливість харчуватися. Спостереження за досліджуваними мухами проводили з інтервалом у три години (вісім разів на добу). При цьому визначалася кількість особин, що вижили. В перші дні досліду смертність мух ще не дуже висока, пізніше починається масова їх загибель. Підрахунок вели до моменту загибелі половини мух у пробірці (Lt50). Тривалість життя мух за умов голодування виражали у годинах. Досліди проводили в десятикратній повторності.
Активність АДГ в экстрактах тканин дрозофіли визначали спектрофотометрично на СФ-26, використовуючи нкубаційну суміш наступного складу: 0,05 М тріс-НCL буфер, pH 8,5; 1,2 мM NAD+; 0,1 M спирт. Активність ферменту оцінювали по збільшенню вмісту NADН в середовищі інкубації на протязі 3 хвилин. В якості субстрату використовували ізопропанол. Активність АДГ виражали в нмолях NADН/хв∙мг білку [McKechnie, Geer, 1984].
Статистичну обробку отриманих даних здійснювали за допомогою критерія Стьюдента (Рокицкий, 1973)
1). Визначали середнє значення тривалості життя в умовах голодування:
![]() , де
, де
М – середнє значення тривалості життя при голодуванні; n кількість дослідів.
2). Визначали квадратичне відхилення:
![]() , де
, де
М – а – різниця середнього та індивідуальних значень тривалост життя.
3). Визначали помилку середньоарифметичного значення тривалост життя:
![]() , де
, де
m – помилка середньоарифметичного значення; – квадратичне відхилення
При р > 0.05 відмінності не достовірні;
Якщо р < 0,05 – відмінності достовірні
3. Результати досліджень та їх обговорення
дані, отримані в експерименті при вивченн стійкості мух до голодування, наведено в таблиці.
Таблиця
Тривалість життя при голодуванні мух із досліджуваних популяцій, години (Lt50)
| Популяції | Самки | Самці | Самки та самці |
|
Одеса |
39,60 1,60# |
34,50 1,02* # |
37,05 1,55# |
|
Київ |
24,60 2,53** | 23,40 2,06** | 24,00 2,21** |
|
Варва |
32,70 ± 1,22** # |
26,70 ± 1,64* ** |
29,70 ± 1,02** # |
|
Пирятин |
34,80 ± 1,28** # |
29,70 ± 1,26* ** # |
32,25 ± 1,02** # |
|
Озеро |
30,90 ± 1,09** # |
34,50 ± 1,03# |
32,70 ± 1,18** # |
|
Сад |
33,00 ± 1,38** # |
29,40 ± 1,25** # |
31,20 ± 0,10** # |
* – Різниця достовірна порівняно з самками.
** – Різниця достовірна порівняно з Одеською популяцією.
# – Різниця достовірна порівняно з Київською популяцією.
Із представлених в таблиці та на рис. 1 даних видно, що самки самці із популяції Києва не відрізняються по стійкості до голодування, адже тривалість їх життя в умовах досліду приблизно однакова (24,6 2,53 та 23,4 2,06 години відповідно). Так само не виявляють статевих відмінностей за цією ознакою і мухи із Чорнобильської зони. Самки і самці із популяції Озера виживали за голодування 30,90 ± 1,09 і 34,50 ± 1,03 годин відповідно, а із популяції Саду – 33,00 ± 1,38 і 29,40 ± 1,25 годин відповідно. Самки із популяцій Одеси, Варви і Пирятина виявилися більш витривалими в умовах голодування порівняно з самцями.
Години(Lt50)
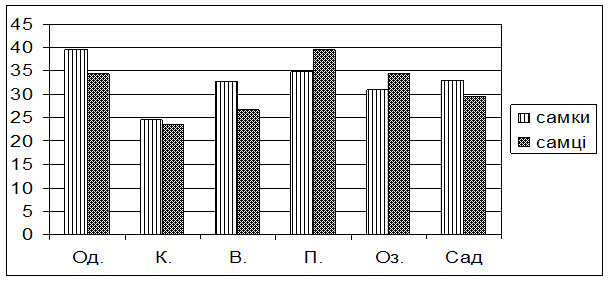
Рис. 1. Тривалість життя при голодуванні самок та самців досліджуваних популяцій дрозофіли
Порівнюючи результати власного експерименту із даними, опублікованими в опрацьованій літературі [Билева, Зимина и др., 1978; Хаустова, Тоцкий, 1990; Хаустова, Тоцкий и др., 2004), можна підтвердити, що самки мають вищу стійкість до голодування, ніж самці, але ця залежність виявляється не завжди і лише за певних умов..
На рис. 2 показані міжпопуляційні відмінності у самок і самців досліджуваних популяцій.
Години (Lt50)
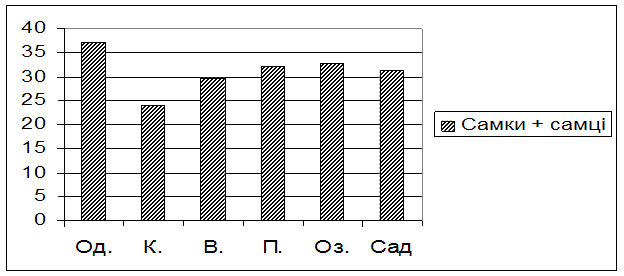
Рис. 2. Тривалість життя в умовах голодування мух із досліджуваних популяцій
Мухи із популяції Одеси виявились найбільш стійкими до голодування – вони прожили 37,05 1,55 годин в даних умовах. Дещо нижчий показник тривалості життя в умовах голодування належить мухам із популяцій Пирятина та Озера і складає 32,25 ± 1,02 та 32,70 ± 1,18 годин відповідно. Досліджувані мухи із популяції Варви і саду мають середні показники тривалості життя, а саме: 29,70 ± 1,02 та 31,20 ± 0,10 години. Найменшу тривалість життя показали мухи, відібрані із популяції Києва, – вони прожили лише 24,00 2,21 годин в дослідних умовах.
Таким чином, мухи із популяції Одеси в цілому мають найвищу стійкість до голодування, а у мух київської популяції даний показник пристосованості виявився найнижчим.
Відмінності по тривалості життя в умовах голодування у досліджуваних популяцій дрозофіли можна пояснити, скоріш за все, наявністю у них різних алельних генів, що вносять різний вклад у генетичну детермінацію даної ознаки. Це також може бути пов’язано з географічним розташуванням досліджуваних природних популяцій.
На рис. 3 представлені дані по активності АДГ у мух досліджуваних популяцій.
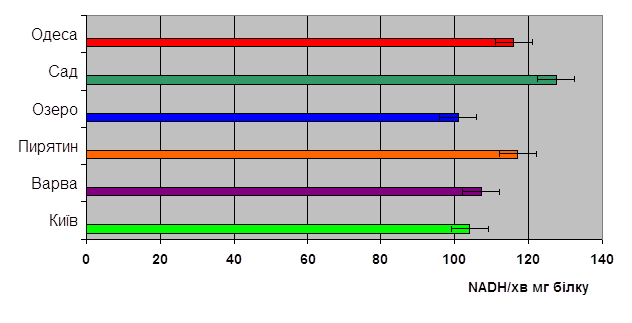 Рис. 3. Активність АДГ
у мух досліджуваних популяцій
Рис. 3. Активність АДГ
у мух досліджуваних популяцій
Аналізуючи дані, приведені на рис. 3, приходимо до висновку, що у мух досліджуваних популяцій є суттєві відмінності в активності АДГ.
Так, найвища активність ферменту (127.46 NADН/хв∙мг білку) спостерігалась у мух з популяції Саду Чорнобильської зони. Високою активністю АДГ відрізнялись мухи із популяцій Пирятина та Одеси (117.02 та 115.97 NADН/хв∙мг білку відповідно). Достовірно нижча активність АДГ порівняно із такою у мух Саду Чорнобильської зони спостерігалась у мух з популяцій Варви та Києва (107.21 та 104.06 NADН/хв∙мг білку). найнижча активність АДГ притаманна мухам озерної популяції (100.93 NADН/хв∙мг білку).
Зазвичай рівень активності АДГ залежить від алельного стану структурного гена Adh. За даними літератури [Chambers, Wilks et all., 1984] електрофоретично швидкорухомий АДГ-F алозим відрізняється високою активністю, а менш рухомий АДГ-S алозим – низькою активністю, але більшою, ніж АДГ-F, термостабільністю ферменту.
в той же час відома залежність деяких показників пристосованості дрозофіли від приналежності мух до того чи іншого Adh-варіанту і від рівня активност ферменту [Хаустова, Тоцкий, 1990]. Установлено, що активність AДГ впливає на виживаність дрозофіли при помірно високій концентрації спирту в середовищі, в умовах голодування, в процесі старіння мух і ін. Причиною цього є надзвичайно важлива роль AДГ в утилізації и детоксикації спиртів – природного середовища снування дрозофіли.
В умовах проведеного експерименту не виявлено прямої залежності між витривалістю мух в умовах голодування і активністю їх АДГ, в той же час аналіз приведених даних (рис. 2, 3) свідчить, що стійким до голодування мухам із Одеської популяції притаманна досить висока активність АДГ, а чутливим до голодування мухам Київської популяції – менша активність ферменту.
Узагальнення
Однією із показових характеристик пристосованості є стійкість до голодування здатність вижити в екстремальних умовах нестачі корму.
Більшість тварин різних видів на протязі життя переживають періоди скорочення раціону і тому у них еволюційно закріплені адаптації, що підсилюють х стійкість до голодування. Знання про генетичні та фізіологічні засади цих адаптацій, їх еволюційні зв`язки (як внутрішньовидові, так і популяційні) отримані значною мірою завдяки дослідженням на дрозофілі.
Так як і інші фенотипічні характеристики пристосованості, стійкість до голодування в значній мірі визначається генотипом, контролюється великою кількістю взаємодіючих генів і проявляє значну мінливість в природних популяціях. Ось чому генотипічно різним особинам зазвичай притаманна неоднакова стійкість до голодування.
В контексті вивчення механізмів адаптації генотипів та популяцій до дії екологічних факторів вважається доцільним вияснити роль ферменту алкгольдегідрогенази (АДГ) в життєдіяльності та адаптації у Drosophila melanogaster. Ген-ензимна система АДГ на протязі тривалого часу притягу увагу численних дослідників в різних областях генетики – від молекулярної до популяційної, завдяки відносно простій ідентифікації ферменту, значному поліморфізму і тій ключовій ролі, що АДГ відіграє в життєдіяльності дрозофіли.
З метою визначення тривалості життя при голодуванні в даній робот дослідження проводили на мухах із популяцій різного походження – К (Київ), Од (Одеса), В (Варва), П (Пирятин), Оз (Озеро із Чорнобильської зони) та сад (Яблуневий сад із Чорнобильської зони).
В результаті проведених досліджень встановлено, що найбільш витривалими до дії голоду виявились самки із популяцій Одеси, Варви Пирятина в порівнянні із самцями цих же популяцій. В цілому, найбільшу стійкість до голодування мають мухи Одеської популяції, а найменшу мухи Київської популяції.
У мух досліджуваних популяцій є суттєві відмінності в активності АДГ. Так, найвища активність ферменту спостерігалась у мух із популяції Саду Чорнобильської зони. Високою активністю АДГ відрізнялись і мухи із популяцій Пирятина та Одеси. Достовірно нижча активність АДГ порівняно із такою у мух Саду Чорнобильської зони спостерігалась у мух із популяцій Варви та Києва. найнижча активність АДГ притаманна мухам озерної популяції.
Підсумовуючи дані проведеного експерименту, стверджуємо, що прямої залежності між тривалістю життя мух в умовах голодування і активністю їх АДГ не виявлено. Однак, аналіз отриманих результатів свідчить, що стійким до голодування мухам із Одеської популяції притаманна досить висока активність АДГ, а чутливим до голодування мухам Київської популяції – менша активність цього ферменту.
Висновки
1. Самки з популяцій Одеси, Варви і Пирятина були більш витривалими в умовах голодування порівняно з самцями.
2. Найбільшу стійкість до голодування виявили мухи Одеської популяції, а найменшу мухи Київської популяції.
3. стійким до голодування мухам із Одеської популяції притаманна досить висока активність АДГ, а чутливим до голодування мухам Київської популяції – менша активність ферменту.
4. В цілому в умовах проведеного експерименту не виявлено прямо залежності між витривалістю мух в умовах голодування і активністю їх АДГ.
Список цитованої літератури
1. Бей-Биенко Г.Я. Определитель насекомых Европейской части СССР - Л: Наука, 1970 – С.199 – 209.
2. Билева Д.С., Зимина Л.Н., Малиновский А.А. Влияние генотипа и среды на продолжительность жизни D. melanogaster // Генетика, 1978. - Т.11 .-№ 5. – С. 848 – 852.
3. Гаврилов Л.А., Гаврилова Н.С. Биология продолжительности жизни М.: Наука, 1991 - 280 с.
4. Лобашев М.Е., Генетика, ЛГУ, 1967 - 718 с.
5. Медведев Н.Н. Практическая генетика - М.: Наука, 1966. - С. 21, 41, 167 - 174.
6. Рокицкий П.Ф. Введение в статистическую генетику.- Минск: Изд-во «Высшая школа», 1973. - 447 с.
7. Тоцький В.М. Генетика. – Одеса: Астропринт, 2008. – 710 с.
8. Тоцкий В.Н., Хаустова Н.Д., Стрельцова Н.А. Полиморфизм алкогольдегидрогеназы и генотипическая адаптация D. melanogaster к действию селективных факторов // Цитология и генетика. – 1995. – Т. 29. – № 6. – С. 54 – 60.
9. Хаустова Н.Д., Тоцкий В.Н. Алкогольдегидрогеназа и адаптация к этанолу у дрозофилы // Генетика. – 1990. – Т. 26. – № 8. – С. 1427 – 1434.
10. Хаустова Н.Д., Моргун С.В. Ген-энзимная система АДГ и приспособленность мутантов D. Melanogaster // Генетика. - 1999 - Т. 35.- № 5. - С. 600 - 605.
11. Хаустова Н.Д. Локус Adh Drosophila melanogaster в условиях отбора на задержку старения // Генетика. – 1995. – Т. 31. – № 5. – С. 646 – 651.
12. Хаустова Н.Д., Тоцкий В.Н., Нассер М.Ю., Стрельцова Н.А. Частота рекомбинаций и приспособленность дрозофилы при структурных перестройках генотипа // Вісник Одеського національного університету. – 2004. – Т. 9. – Вип. 1. – С. 142 – 151.
13. Хвостова В.В., Корочкин Л.И., Голубовский М.Д. Проблемы генетики в исследованиях на дрозофиле. Новосибирск: Наука, 1977. – 282 с.
14. Черник Я.И., Корытко О.Р., Белоконь Е.М. Возрастные изменения алкогольдегидрогеназы, малатдегидрогеназы и лактатдегидрогеназы Drosophila melanogaster // Онтогенез. – 1982. – Т. 13. 5. – С. 169 – 174.
15. Яблоков А.В., Юсуфов А.Г. Эволюционное учение (Дарвинизм). – М.: Высшая школа, 1989. – 335 с.
16. Archer M.A., Phelan, J.P., Beckman, K.A., Rose, M.R. Breakdown in correlations during laboratory evolution. II. Selection on stress resistance in Drosophila populations // Evolution. - 2003. - № 57. - P. 536 – 543.
17. Arsham A.M., Neufeld T.P. Thinking globally and acting locally with TOR. // Curr. Opin. Cell. Biol. - 2006. - № 18. - P. 589 - 597.
18. Burger J.S.M., Hwangbo D.S., Corby-Harris V., Promislow, D.E.L. The functional costs and benefits of dietary restriction in Drosophila // Aging Cell. - 2007. - № 6. - P. 63 - 71.
19. Chambers G.K., Wilks A.V., Gibson J.B. Variation in the biochemical properties of the Drosophila alcohol dehydrogenase allozymes // Biochem. Genet. – 1984. – Vol. 22. – № 1 – 2. – P. 153 – 168.
20. Chippindale A.K., Leroi A.M., Kim S.B., Rose M.R. Phenotypic plasticity and selection in Drosophila life history evolution. 1. Nutrition and the cost of reproduction.// J. Evol. Biol. - 1993. - № 6. - P. 171 - 193.
21. Coyne J.A., Elwyn S. Does the desaturase-2 locus in Drosophila melanogaster cause adaptation and sexual isolation? // Evolution. - 2006. - № 60. - P. 279 - 291.
22. Dorado D., Barbancho M. Differential responses in Drosophila melanogaster to environmental ethanol: modification of fitness components at Adh locus // Heredity. – 1984. – Vol. 53. – № 2. – P. 309 – 320.
23. Greenberg A.J., Moran J.R., Coyne J.A. , Wu C.I. Ecological adaptation during incipient speciation revealed by precise gene replacement. // Science. - 2003. - № 302. - P. 1754 - 1757.
24. Harbison S.T., Yamamoto A.H., Fanara J.J., Norga K.K., Mackay T.F.C. Quantitative trait loci affecting starvation resistance in Drosophila melanogaster. // Genetics. - 2004. - № 166. - Р. 1807 - 1823.
25. Harshman L.G., Moore K.M., Sty M.A., Magwire, M.M. Stress resistance and longevity in selected lines of Drosophila melanogaster. // Neurobiol. Aging. - 1999b. - 20. - P. 521 - 529.
26. Harshman L.G., Schmid J.L. Evolution of starvation resistance in Drosophila melanogaster: aspects of metabolism and counter-impact selection. // Evolution. - 1998. - № 52. - P. 1679 – 1685.
27. Harshman L.G., Hoffmann A.A., Clark A.G. Selection for starvation resistance in Drosophila melanogaster: physiologicalcorrelates, enzyme activities and multiple stress responses. // J. Evol. Biol. - 1999a. - 12. - P. 370 - 379.
28. Huey R.B., Suess J., Hamilton H., Gilchrist G.W. Starvation resistance in Drosophila melanogaster: testing for a possible ’cannibalism’ bias. // Funct. Ecol. - 2004. - № 1. - P. 952 - 954.
29. Kapahi P., Zid B.M., Harpe, T., Koslover D., Sapin V., Benzer, S. Regulation of lifespan in Drosophila by modulation of genes in the TOR signaling pathway. // Curr. Biol. - 2004. - 14. - P. 885 - 890.
30. Leroi A.M., Kim S.B., Rose M.R. The evolution of phenotypic life-history trade-offs – an experimental study using Drosophila melanogaster. // Am. Nat. - 1994. - № 144. - P. 661 - 676.
31. Lin Y.J., Seroude L., Benzer S. Extended life-span and stress resistance in the Drosophila mutant methuselah. // Science. - 1998. - № 282. - P. 943 - 946.
32. Mackay T.F.C., Fry J.D. Polygenic mutation in Drosophila melanogaster: genetic interactions between selection lines and candidate quantitative trait loci. // Genetics. - 1996. - 144. - P. 671 - 688.
33. Partridge L., Piper M.D.W., Mair W. Dietary restriction in Drosophila. // Mech. Ageing Dev. - 2005. - № 126. - P. 938 - 950.
34. Piper M.D.W., Skorupa D., Partridge, L. Diet, metabolism and lifespan in Drosophila. // Exp. Gerontol. - 2005. - № 40. - P. 857 - 862.
35. Piper,M.D.W., Skorupa D., Partridge L. Diet, metabolism and lifespan in Drosophila. // Exp. Gerontol. – 2005. - № 40. - P. 857 - 862.
36. Schmidt P.S., Matzkin L., Ippolito M., Eanes W.F. Geographic variation in diapause incidence, life-history traits, and climatic adaptation in Drosophila melanogaster. // Evolution. - 2005a. - № 59. - P. 1721 - 1732.
37. Tatar M., Bartke A., Antebi A. The endocrine regulation of aging by insulin-like signals. // Science. - 2003. - № 299. - P. 1346 - 1351.
© 2010 Интернет База Рефератов